Articles
- Page Path
- HOME > Restor Dent Endod > Volume 27(5); 2002 > Article
- Original Article Distribution of ion channels in trigeminal ganglion neuron of rat
- Ae-Kyung Kim, Kyoung-kyu Choi, Ho-Young Choi
-
2002;27(5):-462.
DOI: https://doi.org/10.5395/JKACD.2002.27.5.451
Published online: September 30, 2002
Department of Conservative Dentistry, College of Dentistry, Kyung Hee University, Korea.
Copyright © 2002 Korean Academy of Conservative Dentistry
- 1,823 Views
- 2 Download
I. INTRODUCTION
Dorsal root ganglia (DRG) neurons transmit various sensory information such as touch, pressure, pain and temperature from the peripheral region to the central nervous system. Similarly, a pair of trigeminal root ganglia (TRG) is responsible for the sensory inputs from the oromaxillofacial region to which the trigeminal nerve is innervated.
The sensory neurons of mammalian DRG are known to express voltage-dependent ion channels: at least two types of sodium currents (INa) which are distinguished by their sensitivity to TTX (Bossu and Feltz 1984; Caffrey et al. 1992; Elliott and Elliott 1993; Fedulova et al. 1991; Kostyuk et al. 1981, McLean et al. 1988; Ogata and Tatebayashi 1993; Ogata and Tatebayashi 1992a, b; Roy and Narahashi 1992; Schwartz et al. 1990). In addition, the distribution of these ion channels in DRG neurons, according to the size of cells and developmental stages, has been widely studied to elucidate the developmental and functional role of the each type of ion channels in the modulation of specific sensory modalities (Roy and Narahashi 1992; Schwartz et al. 1990).
Voltage-activated calcium channels have been implicated in a number of important neuronal functions, including secretion of neurotransmitters and hormones (Perney et al. 1986; Hirning et al. 1988; Holz et al. 1988), generation of epileptiform discharges (Wong and Prince 1978) and activation of intracellular enzymes (Nishizuka 1984). Ca2+ influx through voltage-activated calcium channels might also contribute to neurodegeneration associated with ischaemia (Choi 1988). For these reasons, considerable attention has been given to defining the criteria for separating different types of Ca2+ channel and for attributing specific functions to the different channels. Although there is general agreement about some properties of Ca2+ channels, it has become clear that more detail information is needed due to an apparent wide diversity in different cell types. In particular, there is little quantitative information on the pharmacological sensitivity of the channel blockers may be useful for distinguishing channel types that are not easily separated on the basis of kinetic properties and may, therefore, be useful in studying the relative contributions of each channel type to specific functions.
All excitable cells express voltage-gated K+ (Kv) channels. These channels are critical for a wide variety of process, including action potential preparation and lymphocyte activation, and play an essential role in neurons where they regulate the resting membrane potential, impact dendritic excitability, control the frequency and duration of action potentials, and modulate neurotransmitter release (Hille 1992).
Several types of inwardly rectifying K+ (Kir) channels have been electrophysiologically identified in retinal Moeller cells (Brew et al. 1986; Ishii et al 1997; Kusaka and Puro 1997; Newman 1993), oligodendrocytes (McLarnon and Kim 1989), glioma cells (Brismar and Collins 1989).
Ca2+-activated K+ channels are present in a remarkable variety of animal cells, where they integrate changes in intracellular Ca2+ concentration with changes in membrane potential. They are involved in many physiological processes including regulation of secretion, smooth muscle tone, and control of action potential shape and firing pattern in excitable cells (Marty 1989; Vergara et al. 1998).
The main goals of this study were (1) to observe some kinds of ion channels (2) to observe the localized distribution of the ion channels in the trigeminal root ganglion neurons that are responsible for the generation and transmission of sensory impulses from oromaxillofacial region to the central nervous system.
II. MATERIALS & METHODS
Rat TRG neurons were prepared by the method described by Liu and Simon (1996) as follows: after the decapitation of the ketamine-anesthesized (100~125mg/kg, I. M.) adult Sprague-Dawley rat (100~150g), a pair of trigeminal root ganglia were dissected and washed several times in cold (4℃) modified Hanks'balanced salt solution (HBSS) containing 130 NaCl, 5 KCl, 0.3 KH2PO4, 4 NaHCO3, 0.3 Na2HPO4, 5.6 D-glucose and 10 N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid (HEPES) (in mM, pH 7.3). They were incubated for 40minutes at 37℃ in HBSS containing 1 mg/ml of type XI collagenase, triturated with flamed Pasteur pipettes, and finally incubated at 37℃ for 6min with 1mg/ml of type IV DNase I. Then they were retriturated and washed/centrifuged three times in Dulbecco's modified Eagle medium (DMEM), and maintained in room temperature.
Isolated trigeminal ganglion neurons were washed complete saline solution, twice, 1min each. The cells were fixed for 10min 4% paraformaldehyde in 0.1M PBS. after three times washing for 10min in 0.1M PBS, the cells incubated for 15min in PBS containing 20% normal goat serum, 1% bovine serum albumin, and 0.1% Triton X-100. Polyclonal anti-Kv 4.2, BKCa, Kir 2.1 α1A, α1B, αE (product no. APC-023, 021, 026 and ACC-001, 002, 006 ; Alomone Labs, Jerusalem, Israel) were used as primary antibodies at a dilution of 1:100 for 24 h at 4℃. The cells were washed for 5 min six times. And then the cells were incubated for 1h at room temperature in PBS containing anti-rabbit fluoresceinlabeled IgG (dilution of 1: 100, KPL Europe). After three 10-min washes in PBS and examined with epifluorescence microscopy.
III. RESULTS
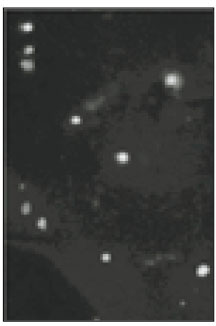
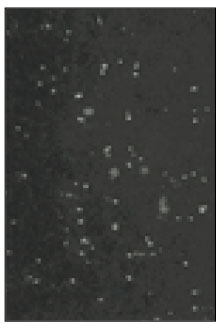
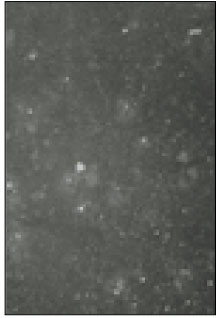
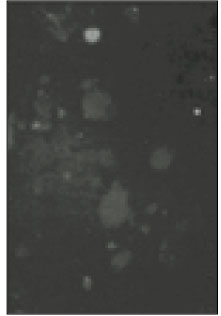
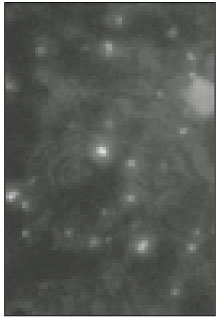
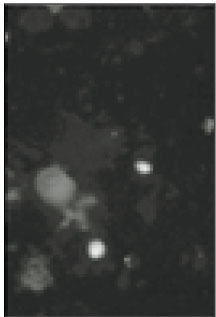
Soma isolated from the trigeminal ganglion were moderately stained with the antibody for the sodium channels. The primary antibodies used in this experiment are able to bind to TTX-s and TTX-r sodium channels. Therefore TTX-s and TTX-r sodium channels on the membrane of TRG neurons could not be separated from each other. Fig. 1 shows TRG neurons observed under fluorescence microscope and Fig. 2 is the micrograph observed in the same field with Fig. 1.
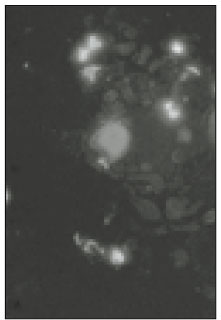
N-type and P/Q-type calcium channels on the TRG neurons were observed under fluorescence microscope (Figs. 3, 5, 7, and 9). Fig. 3 and Fig. 4 show moderately stained trigeminal neurons that treated with N-type calcium channel antibodies, and a light micrograph with the same field. In the magnification of X400 (Fig. 5), only some TRG neurons were stained. This means that N-type calcium channel would not be expressed in all TRG neurons. And intensity of staining was varied from low to high. With the result of this experiment, the relationship of the cell size and the intensity of staining could not be confirmed. Fig. 7 and 9 show P/Q-type calcium channels of the TRG neurons, and distribution of the calcium channels was not uniform but varied in all neurons. This finding could also be found in figure 3 and 5. In this immunocytochemical study we could observe N-type and P/Q-type calcium channels on the TRG neurons.
Inwardly rectifying potassium channels (Kir 2.1), voltage-gated potassium channels (Kv 4.2) and calcium-activated potassium channels (BKCa) on the TRG neurons were observed under fluorescence and light microscopes (Figs. 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22). All the three types of potassium channels were stained with corresponding antibodies. Therefore they exist in TRG neurons of rat. The intensity of staining and the distribution of the channels were similar to those in the case of the calcium channels.
IV. DISCUSSION
Trigeminal sensory nerves relay mechanical, thermal, chemical and proprioceptive information from craniofacial regions. Trigeminal sensory nerves act at the local level to maintain the integrity of craniofacial tissues through trophic influences and serve the whole animal through reflexes that protect other external sensory (e.g. retina, olfactory epithelium, taste receptors) and internal homeostatic (e.g. respiratory and digestive tracts) systems from damaging environmental changes.
The voltage-gated Na+ channels are responsible for the increase in sodium permeability during the initial rapidly rising phase of the action potential in nerve, neuroendocrine, skeletal muscle, and heart muscle cells. Like most cells, these electrically excitable cells maintain a high intracellular K+ concentration and a low intracellular Na+ concentration relative to the extracellular fluid through the energy-dependent pumping of these cations by the Na+-K+- ATPase. Excitable cells also maintain a resting membrane potential, inside negative, because their surface membranes are specifically permeable to K+. Electrically excitable cells are distinguished, however, by having voltage-sensitive ion channels in their surface membranes that respond to membrane potential changes with large regenerative increases in permeabilities to specific ions on a time scale of milliseconds. Changes in ionic permeability are correctly measured only if the membrane potential of the cell is controlled experimentally (Hodgkin and Huxley 1952). The voltage-sensitive Na+ channel, whose opening causes the upstroke of the nerve action potential, is blocked in practically all neurons by low concentrations of TTX Na+ channels in nociceptive neurons have long been known to be an exception as they are TTX-resistant, a property they share with Na+ channels in cardiac muscle (Elliot and Elliot 1993; Cohen and Barchi 1993). A TTX-resistant Na+ channel has recently been isolated by difference cloning of mRNA from the dorsal root ganglion (DRG) (Akopian et al. 1996).
Classification of primary afferent neurons of the mammalian DRG is based on the morphological and electrophysiological characteristics of neurons (Harper and Lawson 1985a, b). The cell size is known to be correlated with the axonal conduction velocity very well : large neurons with myelinated Aα, β fibers are fast conducting, while small cells with myelinated Aδ and unmyelinated C fibers are slow conducting (Harper and Lawson 1985a). But properties of the somatic action potentials are thought to be associated with the cell types at lesser extent : long action potential with inflected repolarization which is mainly recorded from the Aβ and C cells, and short action potential without plateau during the falling phase recorded in Aα and Aδ cells (Harper and Lawson 1985b). Patch clamp techniques have been used to investigate the range and properties of ionic channels underlying these electrical activity in dissociated mammalian DRG neurons, and two types of INa have been characterized : in general, TTX-s INa are found in large-diameter cells, and TTX-r INa are expressed in small-diameter cells (Caffrey et al. 1992; Elliott and Elliott 1993; Kostyuk et al. 1981; McLean 1988; Ogata and Tatebayashi 1992a; Puil et al. 1986; Schwartz et al. 1990). Though it is generally accepted that TTX-r INa is found in the soma of small DRG neurons that give rise, primarily, to C-type afferent fibers, the frequency of recording TTX-r INa in small neurons is in disagreement among studies from the same type of cell. For example, Caffrey et al. (1992) reported that TTX-r INa was always observed in conjunction with TTX-s INa in small DRG neurons from adult rats, where as Elliott and Elliott (1993) suggested that only 45% of small cells express TTX-r INa from the same preparation. In this experiment, neurons isolated from TRG were 16 to 40µm in diameter. Because primary antibodies used in this experiment are able to bind TTX-s Na+ channels and TTX-r Na+ channels, positive staining of this result shows the presence of both TTX-s Na+ channels and TTX-r Na+ channels in the TRG neurons. And this result shows that the degree of staining does not related with the size of the neuron. Calcium influx, which is mediated by the opening of voltage-dependent calcium channels, triggers a number of cellular processes, including muscle contraction, second-messenger activation cascade, regulation of axonal guidance, control of neurotransmitter release, and long-term changes in neuronal plasticity (Clapham 1995). Over the past few years, molecular cloning techniques combined with the use of specific toxins have allowed the identification of an unexpected variety of calcium channels, which have been classified into different groups, referred to as T, L, N, P, Q, and R, on the basis of their physiological and pharmacological properties (Tsien et al. 1988; Bean 1989; McCleskey 1994). In most cases, multiple calcium channel types coexist in the same neuron and it has been speculated that they may contribute toward different neuronal functions, possibly depending on their localization (Doughty et al. 1998; Wu et al. 1999).
Previous work on L-type and N-type calcium channels has demonstrated different localizations in specific areas of the neuronal surface. L-type channels have been found in the neuronal somata and proximal dendrites of rat brain, spinal cord, and retina (Westenbroek et al. 1990; Ahlijanian et al. 1990), which is consistent with a role in mediating the increases in intracellular calcium occurring in the cell body in response to excitatory inputs to the dendrites. N-type channels have been detected at the release face of presynaptic nerve terminals at the frog neuromuscular junction (Robitaille et al. 1990; Cohen et al. 1991; Torri Tarelli et al. 1991), in the presynaptic calyx-type nerve terminals of chick ciliary ganglia, and in the medial nucleus of the trapezoid body in rat (Haydon et al. 1994; Wu et al. 1999). These data, together with a number of functional findings (Miller 1990; Dunlap et al. 1995; Tsien et al. 1995; Stanley 1997), clearly indicate that N-type channels play a role in supporting neurotransmitter release. However, it is interesting to note that they have also been found on the somata, throughout the dendritic arbor, and on the dendritic spines of CA1 neurons in hippocampal brain slices, thus suggesting that they may also participate in the generation of calcium conductances in distal dendrites (Mills et al. 1994). In this experiment, N-type calcium channels of the somata of TRG neurons were moderately stained with the specific antibodies. Although N-type calcium channels have a role in supporting neurotransmitter release, this result is similar to data that N-type calcium channels have been found on the somata of hippocampal CA1 neurons.
After the initial description of P-type calcium channels in Purkinje cells (Llinas et al. 1989; Mintz et al. 1992) and the subsequent designation of the Q-type calcium channels as a separate channel category (Randall and Tsien 1995), concern has arisen regarding the distinction between them. P/Q-type calcium channels are widely expressed in the mammalian brain in a predominantly presynaptic distribution (Westenbroek et al. 1995), and have a important role in modulating neurotransmitter release (Regehr and Mintz 1994). In this immunocytochemical study, P/Q-type calcium channels were widely stained on the membranes of the TRG neurons. This means that P/Q-type calcium channels exist not only in the central nervous system but also in the peripheral afferent nerves, and they may be concern with the signal transmission in the primary sensory afferent of the trigeminal nerves.
Inwardly rectifying potassium channels (Kir) control the resting K+ conductance in many excitable and non-excitable cells (Hille 1992). They mediate a high K+ conductance at the K+ reversal potential and at voltages slightly positive to the K+ reversal potential. Thus Kir have a stabilizing effect on the resting potential. Whenever Kir are regulated by second messengers, this represents a highly effect mechanism of controlling excitability (Nilius et al. 1993). Various subtypes of the Kir family have been cloned and heterologously expressed in order to study their functional properties. One of the Kir, Kir 2.1 is widely distributed in various tissues. Kir 2.1 is found in mammalian lens epithelial cells (Cooper et al. 1991), ventricular myocytes (Josephson 1998), endothelial cells (Silver and DeCoursey 1990), and oocytes (Ruppersberg and Fakler 1996). In this experiment, we could also find the Kir 2.1 on the membrane of the TRG neurons. As we expected Kir 2.1 was expressed moderately, however its distribution was not so uniform.
Much interest has been generated recently in the potassium channel subunit Kv 4.2 for its critical role in regulating membrane excitability. Several lines of evidence suggest that Kv 4.2 underlies the A-type current (IA) in neurons (Serodio et al. 1996; Song et al. 1998) as well as the transient outward current (Ito) in cardiac ventricular myocytes (Barry et al. 1998). In particular, Tkatch, et al. (2000) have found that Kv 4.2 mRNA abundance and A-type current amplitude are linearly related in the striatum. In the hippocampus, such A-type current has been found to dampen back-propagation of action potentials in the distal dendrites of CA1 pyramidal neurons, thereby setting up the capacity to modify the excitatory postsynaptic potential or back-propagating action potentials following synaptic activity (Hoffman et al. 1997). Immunohistochemical studies show that Kv 4.2 protein localizes abundantly to the hippocampus (Sheng et al. 1992) with particular localization to the neuronal soma and dendrites (Maletic-Savatic et al. 1995), structures in which modulating membrane excitability could have important implications for long-term potentiation and memory. Furthermore, ultrastructural studies in supraoptic neurons have shown that Kv 4.2 is localized to the postsynaptic membrane directly across from the presynaptic terminal (Alonso and Widmer 1997). There is little study on the distribution of Kv 4.2 in primary sensory neurons. This study is the first one about the localization of Kv 4.2 in TRG neurons. Kv 4.2 was expressed moderately on the membrane of the TRG, however, the intensity of staining does not show an uniform pattern in each neuron. Although this result means that some of the TRG neurons do not have or have some of Kv 4.2, the reason and the physiological implication of the result were not fully understood.
Ca2+-activated K+ channels (KCa)are present in a remarkable variety of animal cells, where they integrate changes in intracellular Ca2+ concentration with changes in membrane potential. They are involved in many physiologic processes including regulation of secretion, smooth muscle tone, and control of action potential shape and firing pattern in excitable cells (Marty 1989; Vergara et al. 1998). KCa can be divided into three main subfamilies based on their electrophysiological, pharmacological, and molecular profiles: largeconductance Ca2+-activated K+ channels, intermediate-conductance Ca2+-activated K+ channels, and small-conductance Ca2+-activated K+ channels (Vergara et al. 1998). Large-conductance Ca2+-activated K+ channels (BKCa) are found in a variety of both electrically excitable and nonexcitable cells (Jan and Jan 1997). Their activities are triggered by membrane depolarization and enhanced by cytosolic Ca2+, providing a link between the metabolic and electrical state of cells. The physiological roles of BKCa channels have been examined in several tissues. In smooth muscle in which they are particularly abundant, BKCa channels play a key role in setting the pace of contractile activity (Nelson et al. 1995), and in neurons, they are involved in regulation of transmitter release and repolarization of action potentials (Kaczorowski et al. 1996). In this experiment BKCa channels were expressed in TRG neurons. Because the BKCa channels regulate the neurotransmitters and are activated after depolarization, they may involve the release the neurotransmitters in trigeminal nerve. The TRG neurons were moderately stained with the specific antibodies for the BKCa channels.
In this immunocytochemical study, thus, we could observe (1) the sodium channels which can be divided into TTX-s and TTX-r sodium channels, (2) the calcium channels: N-type and P/Q-type, and (3) the potassium channels: inwardly rectifying potassium (Kir 2.1) channels, voltage-gated potassium (Kv 4.2) channels, and large-conductance calcium-activated potassium (BKCa) channels in TRG neurons.
V. CONCLUSION
Dorsal root ganglia (DRG) neurons transmit various sensory information such as touch, pressure, pain and temperature from the peripheral region to the central nervous system. Similarly, a pair of trigeminal root ganglia (TRG) is responsible for the sensory inputs from the oromaxillofacial region to which the trigeminal nerve is innervated. Ionic channels in neuronal membranes have a fundamental role in the transmission of various sensory impulses. Particularly, sodium channels are important to generation action potentials and classified into TTX-r and TTX-s sodium channels.
Calcium influx, which is mediated by the opening of voltage-dependent calcium channels, triggers a number of cellular processes, including muscle contraction, second-messenger activation cascade, regulation of axonal guidance, control of neurotransmitter release, and long-term changes in neuronal plasticity.
There are many sets of potassium channels with different subunits. Therefore potassium channels can be divided into several categories. Generally potassium channels are involved in repolarization after depolarization, setting the pace of contractile activity, regulation of neurotransmitter release, and modulating membrane excitability.
The main goals of this study were (1) to observe some kinds of ion channels (2) to observe the localized distribution of the ion channels in the trigeminal root ganglion neurons that are responsible for the generation and transmission of sensory impulses from oromaxillofacial region to the central nervous system.
In this immunocytochemical study.
We observed the sodium channels which can be divided into TTX-s and TTX-r sodium channels in TRG neurons.
We observed the calcium channels: N-type and P/Q-type in TRG neurons.
We observed the potassium channels: inwardly rectifying potassium (Kir 2.1) channels, voltage-gated potassium (Kv 4.2) channels, and large-conductance calciumactivated potassium (BKCa) channels in TRG neurons.
- 1. Ahlijanian MK, Westenbroek RE, Catterall WA. Subunit structure and localization of dihydropyridine-sensitive calcium channels in mammalian btain, spinal cord, and retina. Neuron. 1990;4: 819-832.ArticlePubMed
- 2. Akopian AN, Sivilotti L, Wood JNA. tetrodotoxin-resistant voltage-gated sodium channel expressed by sensory neurons. Nature. 1996;379: 257-262.ArticlePubMedPDF
- 3. Alonso G, Widmer H. Clustering of Kv4.2 potassium channels in Phostsy.
- 4. Barry DM, Xu H, Schuessler RB, Nerbonne JM. Functional knockout of the transient outward current, long-QT syndrome, and cardiac remodeling in mice expressing a dominant-negative Kv4 alpha subunit. Circ Res. 1998;83: 560-567.PubMed
- 5. Bean BP. Classes of calcium channels in vertebrate cells. Annu Rev Physiol. 1989;51: 367-384.ArticlePubMed
- 6. Bignami A. Neuron-Glia Interrelations During Phylogeny. I. Phylogeny and Ontogeny of Glial Cells. 1995;New Jersey: Humanna; 3-39.
- 7. Bossu JL, Feltz A. Patch-clamp study of the tetrodotoxin-resistant sodium current in group C sensory neurones. Neurosci Lett. 1984;51: 241-246.ArticlePubMed
- 8. Brew H, Gray PT, Mobbs P. Endfeet of retinal glial cells have higher densities of ion channels that mediate K+ buffering. Nature. 1986;324: 466-468.ArticlePubMedPDF
- 9. Brismar T, Collins VP. Inward rectifying potassium channels in human malignant glioma cells. Brain Res. 1989;480: 249-258.ArticlePubMed
- 10. Caffrey JM, Eng DL, Black JA, Waxman SG, Kocsis JD. Three types of sodium channels in adult rat dorsal root ganglion neurons. Brain Res. 1992;592: 283-297.ArticlePubMed
- 11. Choi DW. Calcium-mediated neurotoxicity : relationship to specific channel types and role in ischemic damage. Trends Neurosci. 1988;11: 465-469.ArticlePubMed
- 12. Clapham DE. Calcium signaling. Cell. 2007;131: 1047-1058.ArticlePubMed
- 13. Cohen SA, Barchi RL. Voltage-dependent sodium channels. Int Rev Cytol. 1993;137: 55-103.
- 14. Cohen MW, Jones OT, Angelides KJ. Distribution of Ca2+ channels on frog motor nerve terminals revealed by fluorescent ω-conotoxin. J Neurosci. 1991;11: 1032-1039 Cooper, K., Rae, J. L., AND Deway, J. Inwardly rectifying potassium current in mammalian lens epithelial cells. Am J Physiol. 261 : C115-23.ArticlePubMedPMC
- 15. Doughty JM, Barnes-Davies M, Rusznak Z, Harasztosi C, Forsythe ID. Constructing Ca2+ channel subtypes at cell bodies and synaptic terminals of rat anterioventral cochlear bushy neurons. J Physiol. 1998;512: 365-376 Dunlap, K., Luebke, J. I., AND Turner, T. J. Exocytotic Ca2+ channels in mammalian central neurons. Trends Neurosci. 18 : 89-98, 1995.PubMedPMC
- 16. Elliott AA, Elliott JR. Characterization of TTX-sensitive and TTX-resistant sodium currents in small cells from adult rat dorsal root ganglia. J Physiol. 1993;463: 39-56.ArticlePubMedPMC
- 17. Fedulova SA, Kostyuk PG, Veselovsky NS. Ionic mechanisms of electrical excitability in rat sensory neurons during postnatal ontogenesis. Neuroscience. 1991;41: 303-309.ArticlePubMed
- 18. Harper AA, Lawson SN. Conduction velocity is related to morphological cell type in rat dorsal root ganglion neurones. J Physiol. 1985a;359: 31-46.Article
- 19. Harper AA, Lawson SN. Electrical properties of rat dorsal root ganglion neurons with different peripheral nerve conduction velocities. J Physiol. 1985b;359: 47-63.
- 20. Haydon PG, Henderson E, Stanley E. Localization of individual calcium channels at the release face of a presynaptic nerve terminal. Neuron. 1994;13: 1275-1280.ArticlePubMed
- 21. Hille B. Ionic channels of Excitable Membranes. 1992;Sunderland, MA: Sinauer.
- 22. Hirning LD, Fox AP, McCleskey EW, Olivera BM, Thayer SA, Miller RJ, Tsien RW. Dominant role of N-type Ca2+ channels in evoked release of norepinephrine from sympathetic neurons. Science. 1988;239: 57-61.ArticlePubMed
- 23. Hodgkin AL, Huxley AF. Currents carried by sodium and potassium ions through the membrane of the giant axon of Loligo. J Physiol. 1952;116: 449-472.ArticlePubMedPMCPDF
- 24. Hoffman DA, Magee JC, Colbert CM, Johnston D. K+ channel regulation of signal propagation in dendrites of hippocampal pyramidal neurons. Nature. 1997;387: 869-875.ArticlePubMedPDF
- 25. Holz GG, Dunlap K, Kream RM. Characterization of the electrically evoked release of substance P from dorsal root ganglion neurons : methods and dihydropyridine sensitivity. J Neurosci. 1988;8: 463-471.ArticlePubMedPMC
- 26. Ishii M, Horio Y, Tada Y, Hibino H, Inanobe A, Ito M, Yamada M, Gotow T, Uchiyama Y, Kurachi Y. Expression and clustered distribution of an inwardly rectifying potassium channel, KAB-2/Kir4.1, on mammalian retinal Muller cell membrane: the regulation by insulin and laminin signals. J Neurosci. 1997;17: 7725-7735.ArticlePubMedPMC
- 27. Jan LY, Jan YN. Cloned potassium channels from eukaryotes and prokaryotes. Annu Rev Neurosci. 1997;20: 91-123.ArticlePubMed
- 28. Josephson IR. Properties of inwardly rectifying K+ channels in venricular myocytes. Mol Cell Biochem. 1988;80: 21-26.PubMed
- 29. Kaczorowski GJ, Knaus HG, Leonard RJ, Mcmanus OB, Garcia ML. High-conductance calcium-activated potassium channels : structure, pharmacology, and function. J Bioenerg Biomembr. 1996;28: 255-267.ArticlePubMedPDF
- 30. Kostyuk PG, Veselovsky NS, Fedulova SA, Tsyndrenko AY. Ionic currents in the somatic membrane of rat dorsal root ganglion neurons - I. Sodium currents. Neuroscience. 1981;6: 2423-2430.ArticlePubMed
- 31. Kusaks S, Puro DG. Intracellular AYP activates inwardly rectifying K+ channels in human and monkey retinal Muller (glial) cells. J Physiol. 1997;500: 593-604.PubMedPMC
- 32. Llinas R, et al. Blocking and isolation of a calcium channel from neurons in mammals and cephalopods utilizing a toxin fraction (FIX) from the funnel web spider poison. Proc Natl Acad Sci USA. 1989;86: 1689-1693.PubMedPMC
- 33. Maletic-Savatic M, Lenn NJ, TRIMMER JS. Differential spatiotemporal expression of K+ channel polypeptides in rat hippocampal neurons developing in situ and in vitro. J Neurosci. 1995;15: 3840-3851.ArticlePubMedPMC
- 34. Marty A. The physiological role of calcium-dependent channels. Trends Neurosci. 1989;12: 420-424.ArticlePubMed
- 35. McCleskey EW. Calcium channels : Celluar roles and molecular mechanisms. Curr Opin Neurobiol. 1994;4: 304-312.PubMed
- 36. Mclarnon JG, Kim SU. Existence of inward potassium currents in adult human oligodendrocytes. Neurosci Lett. 1989;101: 107-112.ArticlePubMed
- 37. McLean MJ, Bennet PB, Thomas RM. Subtypes of dorsal root ganglion neurons based on different inward currents as measured by whole cell voltage clamp. Mol Cell Biochem. 1988;80: 94-107.ArticlePDF
- 38. Miller RJ. Receptor-mediated regulation of calcium channels and neurotransmitter release. FASEB J. 1990;4: 3291-3299.ArticlePubMedPDF
- 39. Mills LR, Niesen CE, So AP, Carlen PL, Spigelman I, Jones OT. N-type Ca2+ channels are located on somata, dendrites and a subpopulation of dendritic spines on live hippocampal pyramidal neurons. J Neurosci. 1994;14: 6815-6824.ArticlePubMedPMC
- 40. Mintz I, et al. P type calcium channels in rat central and peripheral neurons. Neuron. 1992;9: 85-95.ArticlePubMed
- 41. Nelson MT, Cheng H, Rubart M, Santana LF, Bonev AD, Knot HJ, Lederer WJ. Relaxation of arterial smooth muscle by calcium sparks. Science. 1995;270: 633-637.ArticlePubMed
- 42. Newman EA. Inward-rectifying potassium channels in retinal glial (Muller) cells. J Neurosci. 1993;13: 3333-3345.ArticlePubMedPMC
- 43. Nilius B, Schwarz G, Droogmans G. Modulation by histamine of an inwardly rectifying potassium channel in huma endotherial cells. J Physiol. 1993;472: 359-371.PubMedPMC
- 44. Nishizuka Y. The role of protein kinase C in cell surface signal transduction and tumour promotion. Nature. 1984;308: 693-698.ArticlePubMedPDF
- 45. Ogata N, Tatebayashi H. Kinetic analysis of two types of Na+ channels in rat dorsal root ganglia. J Physiol. 1993;466: 9-37.ArticlePubMedPMC
- 46. Ogata N, Tatebayashi H. Ontogenic development of the TTX-sensitive and TTX-insensitive Na+ channels in neurons of the rat dorsal root ganglia. Brain Res Dev Brain Res. 1992a;65: 93-100.Article
- 47. Ogata N, Tatebayashi H. Slow inactivation of tetrodotoxin-insensitive Na+ channels in neurons of rat dorsal root ganglia. J Membr Biol. 1992b;129: 71-80.ArticlePDF
- 48. Perney TM, Hirning LD, Leeman SE, Miller RJ. Multiple calcium channels mediate neurotransmitter release from peripheral neurons. Proc Natl Acad Sci U S A. 1986;83: 6656-6659.ArticlePubMedPMC
- 49. Puil E, Gimbarzevsky B, Miura RM. Quantification of membrane properties of trigeminal root gangion neurons in guinea pigs. J Neurophysiol. 1986;55: 995-1016.PubMed
- 50. Randall A, Tsien R. Pharmacological dissection of multiple types of calcium channel currents in rat cerebellar granule neurons. J Neurosci. 1992;15(4):2995-3012.ArticlePubMedPMC
- 51. Regehr WG, Mintz IM. Participation of multiple calcium channel types in transmission at single climbing fiber to Purkinje cell synapses. Neuron. 1900;12: 605-613.Article
- 52. Robitaille R, Adler EM, Charlton MP. Strategic location of calcium channels at transmitter release sites of frog neuromuscular synapes. Neuron. 1990;5: 773-779.PubMed
- 53. Roy ML, Narahashi T. Differential properties of tetrodotoxin-sensitive and tetrodotoxin -resistant sodium channels in rat dorsal root ganglion neurons. J Neurosci. 1992;12: 2104-2111.ArticlePubMedPMC
- 54. Schwartz A, Palti Y, Meiri H. Structural and developmental differences between three types of Na channels in dorsal root ganglion cells of newborn rats. J Membr Biol. 1990;116: 117-128.ArticlePubMedPDF
- 55. Serodio P, Vega-SaenzDeMiera E, Rudy B. Cloning of a novel component of A-type K+ channels operating at subthreshold potentials with unique expression in heart and brain. J Neurophysiol. 1900;75: 2174-2179.Article
- 56. Sheng M, Tsaur MI, Jan YN. Subcellular segregation of two A-type K+ channel proteins in rat central neurons. Neuron. 1992;2: 271-284.Article
- 57. Silver MR, Decoursey TE. Intinsic gating of inward rectifier in bovine pulmonary artery endothelial cells in the presence or absence of internal Mg2+. J Gen Physiol. 1990;96: 109-133.PubMedPMC
- 58. Song WJ, Tkatch T, Baranauskas G, IChinohe N, Kitai ST, Surmeier DJ. Somatodenditic depolarization-activated potassium currents in rat neostriatal cholinergic interneurons are predominantly of the A type and attributable to coexpression of Kv42 and Kv 41 subunits. J Neurosci. 1998;18: 3124-3137.ArticlePubMedPMC
- 59. Stanley EF. The calcium channel and the organization of the presynaptic transmitter release face. Trends Neurosci. 1997;20: 404-409.ArticlePubMed
- 60. Tkatch T, Baranauskas G, Surmerier DJ. Kv4.2 mRNA abundance and A-type K(+) current amplitude are linearly related in basal ganglia and basal forebrain neurons. J Neurosci. 2000;20: 579-588.ArticlePubMedPMC
- 61. Torri-Tarelli F, Passafaro M, Clementi F, Sher M. Presynaptic localization of omegaconotoxin-sensitive calcium channels at the frog neuromuscular junction. Brain Res. 1991;547: 331-334.PubMed
- 62. Tsien RW, Lipscombe D, Mandison DV, Bley K, Fox A. Reflections on Ca2+ channel diversity, 1984-1994. Trends Neurosci. 1995;18: 52-54.PubMed
- 63. Tsien RW, Lipscomble D, Mandison DV, Bley K, Fox A. Multiple types of neuronal calcium channels and their selective modulation. Trends Neurosci. 1988;11: 431-438.ArticlePubMed
- 64. Vergara C, Latorre R, Marrion NV, Adelman JP. Calcium-activated potassium channels. Curr Opin Neurobiol. 1998;8: 321-329.ArticlePubMed
- 65. Westenbroek RE, Ahlijanian MK, Catterall WA. Clustering of L-type Ca2+ channels at the base of major dendrites in hippocampal pyramidal neurons. Nature. 1990;347: 281-284.ArticlePubMedPDF
- 66. Westenbroek RE, Sakurai T, Elliot EM, et al. Immunochemical identification and subcellular distribution of the alpha 1A subunits of brain calcium channels. J Neurosci. 1995;15: 6403-6418.ArticlePubMedPMC
- 67. Wong RKS, Prince DA. Participation of calcium spikes during intrinsic burst firing in hippocampal neurons. Brain Research. 1978;159: 385-390.ArticlePubMed
- 68. Wu LG, Westenbroek RE, Borst JGG, Catteral WA, Sakmann B. Calcium channel types with distinct presynaptic localization couple differentially to transmitter release in single calyx-type synapses. J Neurosci. 1999;19: 726-736.ArticlePubMedPMC
REFERENCES












Tables & Figures
REFERENCES
Citations
Citations to this article as recorded by 
 ePub Link
ePub Link Cite
CiteDistribution of ion channels in trigeminal ganglion neuron of rat
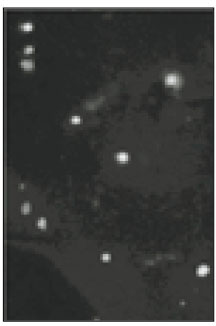

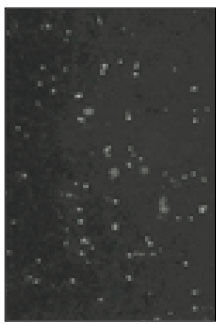

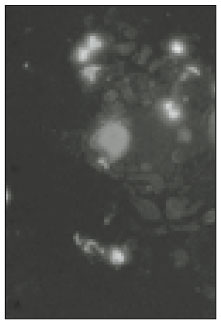

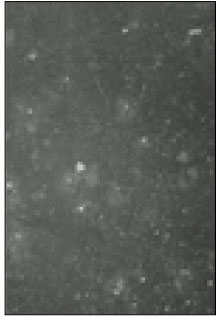

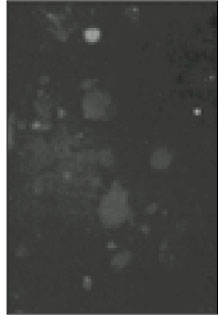








Fig. 1
Fluorescent microscopic image of sodium channels.
Fig. 2
Microscopic image of sodium channels.
Fig. 3
Fluorescent microscopic image of N-type calcium channels. (×100)
Fig. 4
Microscopic image of N-type calcium channel. (×100)
Fig. 5
Fluorescent microscopic image of N-type calcium channels. (×400)
Fig. 6
Microscopic image of N-type calcium channels. (×400)
Fig. 7
Fluorescent microscopic image of P/Q type calcium channels.(×100)
Fig. 8
Microscopic image of P/Q type calcium channels.(×100)
Fig. 9
Fluorescent microscopic image of P/Q type calcium channels.(×400)
Fig. 10
Microscopic image of P/Q type calcium channels. (×400)
Fig. 11
Fluorescent microscopic image of Kir 2.1 potassium channels. (×100)
Fig. 12
Microscopic image of Kir 2.1 potassium channels. (×100)
Fig. 13
Fluorescent microscopic image of Kir 2.1 potassium channels. (×400)
Fig. 14
Microscopic image of Kir 2..1 potassium channels. (×400)
Fig. 15
Fluorescent microscopic image of Kv 4.2 potassium channels. (×100)
Fig. 16
Microscopic image of Kv 4.2 potassium channels. (×100)
Fig. 17
Fluorescent microscopic image of Kv 4.2 potassium channels. (×400)
Fig. 18
Microscopic image of Kv 4.2 potassium channels. (×400)
Fig. 19
Fluorescent microscopic image of BKCa potassium channels. (×100)
Fig. 20
Microscopic image of BKCa potassium channels. (×100)
Fig. 21
Fluorescent microscopic image of BKCa potassium channels. (×400)
Fig. 22
Microscopic image of BKCa potassium channels. (×400)
Fig. 1
Fig. 2
Fig. 3
Fig. 4
Fig. 5
Fig. 6
Fig. 7
Fig. 8
Fig. 9
Fig. 10
Fig. 11
Fig. 12
Fig. 13
Fig. 14
Fig. 15
Fig. 16
Fig. 17
Fig. 18
Fig. 19
Fig. 20
Fig. 21
Fig. 22
Distribution of ion channels in trigeminal ganglion neuron of rat