Articles
- Page Path
- HOME > Restor Dent Endod > Volume 29(5); 2004 > Article
-
Original Article
MMP-1 and TIMP-1 production in MG-63 cells stimulated with
Prevotella nigrescens lipopolysaccharide -
Won-Kyung Yang1, Mi-Ri Kim2, Won-Jun Shon1, In-Bog Lee1, Byeong-Hoon Cho1
 , Chung-Moon Um1, Ho-Hyun Son1
, Chung-Moon Um1, Ho-Hyun Son1 -
2004;29(5):-478.
DOI: https://doi.org/10.5395/JKACD.2004.29.5.470
Published online: September 30, 2004
1Department of Conservative Dentistry, College of Dentistry, Seoul National University, Korea.
2Department of Conservative Dentistry, Asan Medical Center, Korea.
- Corresponding author: Ho-Hyun Son. Department of Conservative Dentistry, College of Dentistry, Seoul National University, 28 Yoengun-dong, Chongro-gu, Seoul, Korea, 110-749. Tel: 82-2-760-1634, Fax: 82-2-760-3859, hhson@snu.ac.kr
Copyright © 2004 Korean Academy of Conservative Dentistry
- 1,501 Views
- 0 Download
Abstract
-
The purpose of this study is to monitor the secretion of matrix metalloproteinase-1 (MMP-1) and tissue inhibitor of metalloproteinase-1 (TIMP-1) produced by human osteosarcoma cell line (MG63) stimulated with Prevotella nigrescens lipopolysaccharides (LPS), and to compare the level of secretion before and after the treatment of calcium hydroxide on P. nigrescens LPS.LPS was extracted and purified from anaerobically cultured P. nigrescens. MG63 cells were stimulated by the LPS (0, 1, 10 µg/ml) or LPS (10 µg/ml) pretreated with 12.5 mg/ml of Ca(OH)2 for 3 days.Total RNA was isolated from the cell, and real-time quantitative polymerase chain reaction (PCR) was performed for quantification of MMP-1 and TIMP-1.The results were as follows.1. MMP-1 mRNA expression at 48 hr was highly increased by stimulation with P. nigrescens LPS. The increase was dose-dependent.2. When stimulated with 1 µg/ml of LPS, TIMP-1 mRNA expression was highly increased at 24 hr and 48 hr. However, TIMP-1 expression was suppressed at higher concentration (10 µg/ml).3. When P. nigrescens LPS was pretreated with Ca(OH)2, MMP-1 and TIMP-1 gene expression was downregulated.The results of this study suggest that transcriptional regulation of MMP-1 and TIMP-1 by P. nigrescens LPS could be one of the important mechanisms in bone resorption of periapical inflammation. The result of calcium hydroxide on MMP-1 and TIMP-1 gene expression suppression shows that calcium hydroxide detoxified bacterial LPS and thus should be used the medication of choice for intracanal dressings in root canal infected with black-pigmented bacteria.
I. INTRODUCTION
Accumulation of bacterial byproduct, such as lipopolysaccharide (LPS) in the root canal system can stimulate the host cells in the periapical tissue. These cells such as neutrophils, macrophages and lymphocytes are activated and release inflammatory mediators to initiate and augment subsequent inflammatory reactions leading to periapical tissue breakdown1).
As is the tissue made up of organized group of cells attached to an extracellular matrix, tissue homeostasis is maintained by coordinating cell growth and proliferation with the production and turnover of the extracellular matrix2). It is conceivable that, since the major component of the organic bone matrix is collagen, collagenolytic enzyme, synthesized by osteoblasts, may also participate in the degradation of the mineralized collagen during bone resorption3).
Matrix metalloproteinases (MMPs) are well known proteolytic enzymes for the degradation of extracellular matrix and this proteolysis is believed to play an essential role in the development and progression of pulpal and periapical inflammation4). MMPs, produced by osteoblast-like cells, are likely to be involved in various steps of the bone resorption process5).
Tissue inhibitor of metalloproteinase (TIMP) is also secreted from the same cells that secret MMPs and block the MMP activity6,7). The inhibitor was found in assoication with bone tissue in vitro and in vivo in immunohistochemial and biochemical studies8,9), and these inhibitors have been known to inhibit bone resorption by maintaining a delicate balance with activities of extracellular matrix-degrading MMP10). Resorption of bone may be the results of complex interactions between metalloproteinases and inhibitors produced by cells of bone, a process that is at least in part regulated by local cytokines including bacterial LPS5). By detecting the balance between the level of MMP and their inhibitor activation, one of the host degradative pathways in the pathogenesis of bacterial-derived pulpal and periapical lesion will be revealed.
Recently, it was demonstrated that black-pigmented Bacteroides (BPB) species play an important role in tissue disintegration of extracellular matrix in pulpal and periapical diseases4,11). The cells of these lesions respond to LPS of the organisms and produce various inflammatory mediators. LPS, localized in the cell wall of Gram-negative bacteria, is known to have a broad spectrum of biochemical and immunochemical activities that can lead to the destruction of host tissue12). There are increasing evidences that bacterial LPS treated with calcium hydroxide intaracanal dressings may have altered biological properties13,14).
Among these BPB, Prevotella nigrescens is the most commonly isolated species from the infected root canals, and thought to be related to endodontic infections15-17). However, there were little information concerning the effects of P. nigrescens LPS on the activation of MMPs produced from osteoblast-like cells, and the influence of calcium hydroxide treatment. Thus, the purpose of this study is to monitor the secretion of MMP and TIMP produced by osteoblastic cells stimulated with P. nigrescens LPS and to compare the level of secretion before and after the treatment of calcium hydroxide on P. nigrescens LPS.
II. MATERIALS AND METHODS
The bacterial strain used was P. nigrescens ATCC33563, and was cultured in brain heart infusion (BHI) broth supplemented with Hemin (10 µg/ml), vitamin K1 (1 µg/ml), and yeast. Bacterial cells were grown anaerobically at 37℃. After sufficient growing, the bacterial cell pellet was obtained by centrifugation at 10,000 × g for 10 min. This pellet was washed with distilled water and then lyophilized. LPS was extracted and purified by the method as described by Eidhin and Mouton in 199318). The lyophilized bacteria was mixed with 1 ml of distilled water and then boiled for 15 min, with 5 min of inverted mixing period. Following centrifugation at 12,000 × g for 5 min, cellular debris was removed, and the supernatant was collected. Then proteinase K (1 mg dissolved in 50 µg/ml water) was added and incubated for 1 h at 60℃. After the tube was boiled for 5 min to precipitate any residual proteinase K, the supernatant was dialyzed against distilled water. After lyophilize the supernatant, the final crude LPS was redissolved to a concentration of 1 mg/ml of distilled water. The purified LPS was confirmed in 12% SDS-PAGE.
Human osteogenic sarcoma cell line (MG63) was obtained from the ATCC (CRL-1427). For the experiment, these cells were plated on 100mm dish in α-MEM supplemented with 1 mM sodium pyruvate, 1 mM non-essential amino acid, 10% FBS, and antibiotics at 37℃ in 5% CO2. After confluency, cells were passed on 6 well plate and stimulated by LPS (0, 1, 10 µg/ml) for various periods of time(8, 12, 24 h). In addition, cells were stimulated with LPS (10 µg/ml) pretreated with 12.5 mg/ml of Ca(OH)2 for 3 days.
For Real-time PCR amplification, optimal primers were designed using Primer express™ software(PE). They are as follows;
MMP-1 :
TIMP-1 :
GAPDH :
After stimulation of cells with LPS and Ca(OH)2 treated LPS for various periods of time, total RNA from the cell was isolated according to the TRIzol Reagent method (Gibco) as described below. The cells were harvested with 0.5 ml of TRIzol reagent and then 100 ml of chloroform was added and inverted mixed. Following centrifugation at 15,000 rpm for 30 min at 4℃, aqueous phase was collected into a fresh tube and mixed with additional 0.25 ml of isopropyl alcohol. After incubation for 10 min on ice, the sample were centrifuged at 15,000 rpm for 10 min at 4℃ to precipitate RNA. The RNA pellet was washed with 1 ml of 75% Ethanol in DEPC and centrifuged at 15,000 rpm for 10 min at 4℃. The isolated RNA was dissolved in DEPC. Extracted total RNA was quantitied using spectrophotometric method to determine the concentration of the primer received. RT reaction mixture containing RT primer, oligo(dT)20 primer, 1 µg of total RNA, Reverse Transcriptase (Invitrogen) was incubated at 42℃ for 50 min, after which the cDNA sample was stored at -20℃. Thermal cycling parameters for RT reactions are as follows; incubation for 10 min at 25℃, RT for 30 min at 48℃, RT inactivation for 5 min at 95℃.
Real-time quantitative PCR was performed by a GeneAmp 5700 sequence detection system(PE, Applied Biosystems) with CYBR Green Master Mix (PE, Applied Biosystems) to assess gene expression. CYBR Green is highly specific to double strand DNA: upon binding to double-strand DNA, it exhibits a large increase in fluorescence. Direct detection of PCR product is monitored by measuring the increase in fluorescence caused by the binding of CYBR Green to double-strand DNA. The reaction mixture (25µL) contains 0.5 µL of the cDNA sample with 0.25 µM of the PCR primer. Two tube method for target genes and control gene (GAPDH) was used in 96 well plate.
The cycle profile was as follows; 1 cycle at 50℃ for 2 min, 1 cycle at 95℃ for I min, 40 cycles at 95℃ for 15 sec, and 60℃ for 1 min. After the last cycle, a dissociation protocol was performed as follows; a hold at 95℃ for 15 sec, a hold at 60℃ for 20 sec, and a slow ramp (20 min) from 60 to 95℃. The dissociation curve was captured during this slow ramp. Experiments were performed in triplicate. The critical threshold cycle (CT) was defined as the cycle at which fluorescence became detectable above the background and was inversely proportional to the logarithm of the initial number of template molecules. A standard curve was plotted for each primer set with CT values obtained from amplification of known quantities of the genomic DNA. By the standard curve of CT vs (logN) starting copy number for all standards, it can be determined the starting copy number of unknowns by interpolation.
III. RESULTS
After stimulated with 0, 1, 10 µg /ml of Prevotella nigrescens LPS at 8, 24 and 48 h, total mRNA of MMP-1 and TIMP-1 were isolated and evaluated by RT-PCR. The threshold cycle was detected and normalized with control gene, GAPDH. The expression of GAPDH was used to compare assay reproducibility utilizing the ratio between raw data and normalized expression as a percent coefficient by employing PCR. The ratio between RNA content and GAPDH of each sample was shown in Table 1 and 2. (Figure 1, 2)
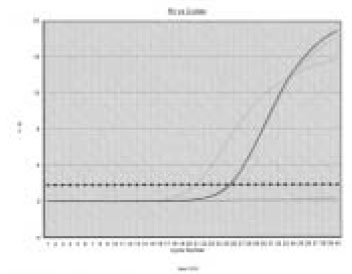
According to the data, MMP-1 gene expression at 48 h was highly increased under the stimulation of both 1 and 10 µg/ml of P. nigrescens LPS. (Table 1, Figure 3). The increase was dose-dependent.
When stimulated with 1 µg/ml of bacterial LPS, TIMP-1 mRNA expression was highly increased at 24 h and 48 h. However, TIMP-1 secretion was suppressed at higher concentration (10 µg/ml) of bacterial LPS (Figure 4).
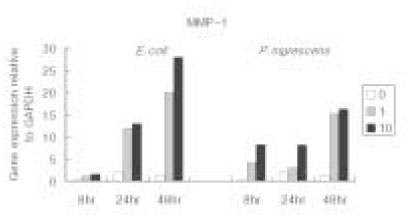
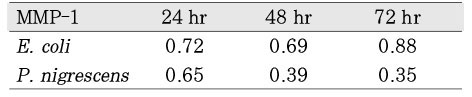
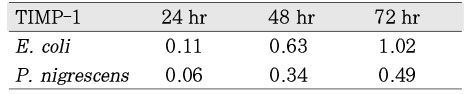
When P. nigrescens LPS was pretreated with Ca(OH)2, MMP-1 and TIMP-1 gene expression was downregulated (Table 3 and 4, Figure 5 and 6).
IV. DISCUSSION
A hallmark of periapical disease is its ability to degrade the surrounding extracellular matrix, resulting in distruction of bone in the periapical tissue. There are increasing evidences that MMPs are essential enzymes responsible for degradation of extracellular matrix in the pathological conditions associated with inflammations19). Numerous studies have examined MMPs secreted from periodontal ligament fibroblast20-22) and correlated the level of expression with the invasive ability of the disease4,6). Individual osteoblasts, like fibroblasts, are capable of both synthesizing and degrading their respective organic matrices in vivo23). MMPs and TIMPs are known to be important in homeostatic matrix remodeling by osteoblasts.
However, MMP and TIMP production under stimulation of bacterial LPS in osteoblastic cell has not been well studied.
Conflicting reports exist concerning the production of MMP-1 by normal human osteoblasts. Rifas et al. reported that human osteoblasts are unable to produce MMP-1 either constitutively or in respons to bone resorptive agents24). De Bart et al. also reported that the mRNA for MMP-1 was not detectable under normal conditions in MG63 cells by Northern blot analysis25).
In the present study, the expression of MMP-1 mRNA was low, but obvious in the negative control. This result consisted with the findings of Panagakos et al. that the expression of MMP-1 was low and essentially invariant over a culture period of human osteoblastic cell line26). This constitutive MMP-1 seems to serve as a remodeling element of connective tissue under physiological conditions27). Otsuka et al. also demonstrated that rat osteosarcoma cell line produced collagenolytic enzymes23).
MMP-1 mRNA was highly expressed in this study after 48 h stimulated with Prevotella nigrescens LPS, and the increase was dose-dependent. The results consisted with many other studies. Nakata et al. demonstrated that the production of MMP-1 was accelerated by sonicated bacterial extracts28). Shin et al. also confirmed this result by detecting high concentration of MMP-1 in relation to clinical symptoms29).
The activity of MMPs is under strict regulation by a group of tissue inhibitor of metalloproteinases(TIMPs), of which TIMP-1 to TIMP-4 have been identified30). TIMP-1, a glycoprotein with a molecular mass of 28kDa, can form a stoichiometric 1:1 complex with MMP-1 and inhibit the activity of MMP-119). Because the same cells that produce MMP-1 are also capable of synthesizing TIMP-1, the overall molar ratio between MMP-1 and TIMP-1 plays a critical role in connective tissue remodeling31).
In this study, TIMP-1 was expressed by MG-63 osteoblastic cells. Otsuka23), Johansen32), Uchida33). also reported that osteoblastic cells synthesized TIMP-1. These results back up our findings.
During the experimental period of time, TIMP-1 expression was highly increased at 24 h and 48 h under the 1 µg/ml of P. nigrescens LPS. However, TIMP-1 secretion was suppressed when exposed to higher concentration (10 µg/ml) of bacterial LPS. The reason of the downregulation of TIMP-1 could be a perturbation in cell-extracellular matrix interaction by the high virulence of bacterial LPS and in turn it culminated in tissue destruction. A better understanding of TIMP activity will be helpful in understanding the etiology of the periapical lesion and its treatment strategies.
LPS is known to exert profound effect on the protein expression by host cells. Wahl et al. has demonstrated that LPS stimulates collagenase release via a mechanism involving PGE2 and cAMP34). This can be the one of the mechanisms that bacterial LPS affect on the development of periapical bone resorption. Prevotella nigrescens LPS was used in this study because infection of the root canals is frequently associated with BPBs and among the isolated black-pigmented bacteroides species, this microorganism is most often isolated from infections of endodontic origin15,17). Gharbia et al. also reported that P. nigrescens is the predominant cultivable BPB from both coronal and apical portions of infected root canals16). E.coli LPS was used as a positive control because its chemical structure and stimulatory effects have been investigated extensively12).
There have been increasing evidences that calcium hydroxide may detoxify bacterial LPS by altering its biological properties13,14,35). Use of calcium hydroxide as a routine intracanal dressing has been advocated in recent years. Calcium hydroxide treatment was effective in suppression of devastating effect of MMP in this study. The fact that the expression of MMP was downregulated when stimulated with either P. nigrescens or control E. coli LPS pretreated with calcium hydroxide indicate that it is effective in preventing extracellular matrix degradation by reducing MMP production from osteoblast. Calcium hydroxide seems to be clinically beneficial when LPS is suspected in the root canal.
IV. CONCLUSION
The mRAN expressions of MMP-1 and TIMP-1 produced by Human osteosarcoma cell line (MG63) stimulated with P. nigrescens LPS (0, 1, 10 µg/ml) and calcium hydroxide, pretreated P. nigrescens LPS (10 µg/ml) for various periods of time, were monitored by real-time PCR.
The results were as follows.
MMP-1 mRNA expression at 48 h was highly increased by stimulation with P. nigrescens LPS. The increase was dose-dependent.
When stimulated with 1 µg/ml of LPS, TIMP-1 mRNA expression was highly increased at 24 h and 48 h. However, TIMP-1 expression was suppressed at higher concentration (10 µg/ml).
When P. nigrescens LPS was pretreated with Ca(OH)2, MMP-1 and TIMP-1 gene expression was downregulated.
The results of this study are suggestive that transcriptional regulation of MMP-1 and TIMP-1 by P. nigrescens LPS is one of the important mechanisms in bone resorption of periapical inflammation. The result of calcium hydroxide on MMP-1 and TIMP-1 gene expression suppression shows that calcium hydroxide detoxified bacterial LPS and thus should be used as a medication of choice for intracanal dressings in root canal infected with black-pigmented bacteria.
- 1. Trowbridge HO. Immunological aspects of chronic inflammation and repair. J Endod. 1990;16: 54-61.ArticlePubMed
- 2. Yang YY, Tsai HF, Lu SC, Huang YF, Chang YC. Regulation of tissue inhibitors of metalloproteinase-1 gene expression by cytokines in human gingival fiboblasts. J Endod. 2002;28: 803-805.PubMed
- 3. Overall CM, Sodek J. Initial characterization of a neutral metalloproteinase, active of native 3/4 collagen fragments, synthesized by ROS 17/2.8 osteoblastic cells, periodontal fibroblasts, and Identified in gingival crevicular fluid. J Dent Res. 1987;66: 1271-1282.ArticlePubMedPDF
- 4. Chang YC, Lai CC, Yang SF, Chan Y, Hsieh YS. Stimulation of matrix metalloproteinases by black-pigmented bacteroides in human pulp and periodontal ligament cell cultures. J Endod. 2002;28: 90-93.ArticlePubMed
- 5. Van der Zee E, Jansen I, Hoeben K, Beertsen W, Everts V. EGF and IL-1α modulate the release of collagenase, gelatinase and TIMP-1 as well as the release of calcium by rabbit calvarial bone explants. J Periodontal Res. 1998;33: 65-72.ArticlePubMed
- 6. Cury JD, Campbell EJ, Lazarus CJ, Albin RJ, Welgus HG. Selective up-regulation of human alveolar macrophage collagenase production by lipopolysaccharide and comparison to collagenase production by fibroblasts. J Immunol. 1988;141: 4306-4312.ArticlePubMedPDF
- 7. Séguier S, Gogly B, Bodineau A, Godeau G, Brousse N. Is collagen breakdown during periodontitis linked to inflammatory cells and expression of matrix metalloproteinases and tissue inhibitors of metalloproteinases in human gingival tissue? J Periodontol. 2001;72: 1398-1406.ArticlePubMed
- 8. Lin SK, Chiang CP, Hong CY, Lin CP, Lan WH, Hsieh CC, Kuo MYP. Immunolocalization of interstitial collagenase(MMP-1) and tissue inhibitor of metalloproteinase-1(TIMP-1) in radicular cysts. J Oral Pathol Med. 1997;26: 458-463.PubMed
- 9. Everts V, Hoegen K, Beertsen W. The release of tissue inhibitor of metalloproteinases by calvarial bone explants and its immunolicalizaiton. Bone Miner. 1993;22: 43-55.PubMed
- 10. Hill PA, Reynolds JJ, Meikle MC. Inhibition of stimulated bone resorption in vitro by TIMP-1 and TIMP-2. Biochem Biophys Acta. 1993;1177: 71-74.ArticlePubMed
- 11. Hosoya S, Matsushima K. Stimulation of interleukin-1β production of human dental pulp cells by Porphylomonas endodontalis lipopolysaccharides. J Endod. 1997;23: 39-42.PubMed
- 12. Koga T, Nishihara T, Fujiwara T, Nisizawa T, Okahashi N, Noguchi T, Hamada S. Biochemical and immunological properties of lipopolysaccharide(LPS) from Bacteroides gingivalis and comparison with LPS from Escherichia coli. Infect Immun. 1985;47: 638-647.ArticlePubMedPMCPDF
- 13. Safavi KE, Nichols FC. Alteration of biological properties of bacterial lipopolysaccharide by calcium hydroxide treatment. J Endod. 1994;20: 127-129.ArticlePubMed
- 14. Safavi KE, Nichols FC. Effects of calcium hydroxide on bacterial lipopolysaccharide. J Endod. 1993;19: 76-78.PubMed
- 15. Baumgartner JC, Watkins BJ, Bae KS, Xia T. Association of black-pigmented bacteria with endodontic infections. J Endod. 1999;25: 413-415.ArticlePubMed
- 16. Gharbia SE, Haapasalo M, Shah H, et al. Characterization of Prevotella intermedia and Prevotella nigrescens isolates from periodontic and endodontic origin. J Periodontol. 1994;65: 56-61.PubMed
- 17. Bae KS, Baumgartner JC, Shearer TR, David LL. Occurrence of Prevotella nigrescens and Prevotella intermedia in infections of endodontic origin. J Endod. 1997;23: 620-623.ArticlePubMed
- 18. Eidhin DN, Mouton C. A rapid method for preparation of rough and smooth lipopolysaccharides from Bacteroides, Porphylomonas and Prevotella. FEMS Microbiol Lett. 1993;110: 133-138.PubMed
- 19. Birkedal-Hansen H, Moore WBI, Bodden MK, Winsor LJ, Brikedal-Hansen B, DeCarlo A, Engler JA. Matrix metalloproteinases: A review. Crit Rev Oral Biol Med. 1993;4: 197-250.ArticlePubMedPDF
- 20. Alvares O, Klebe R, Grant G, Cochran DL. Growth factor effects on the expression of collagenase and TIMP-1 in periodontal ligament cells. J Periodontol. 1995;66: 552-558.ArticlePubMedPDF
- 21. Bolcato-Bellemin AL, Elkaim R, Abehsera A, Fausser JL, Haikel Y, Tenenbaum H. Expression of mRNAs encoding for α and β integrin subunit, MMPs, and TIMPs in stretched human periodontal ligament and gingival fibroblast. J Dent Res. 2000;79: 1712-1716.ArticlePubMedPDF
- 22. Nakaya H, Osawa G, Iwasaki N, Cochran DL, Kamoi K, Oates TW. Effects of bisphophonate enzymes in human periodontal ligament cells. J Periodontol. 2000;71: 1158-1166.PubMed
- 23. Otsuka K, Sodek J, Limeback H. Synthesis of collagenase and collagenase inhibitors by osteoblast-like cells in culture. Eur J Biochem. 1984;145: 123-129.ArticlePubMed
- 24. Rifas L, Halstead LR, Peck WA. Human osteoblasts in vitro secrete tissue inhibitor of metalloproteinases and gelatinase but not interstitial collagenase as major cellular products. J Clin Invest. 1989;84: 686-694.ArticlePubMedPMC
- 25. de Bart AC, Quax PHA, Lowik CWGM, Verheijen . Regulation of plasminogen activation, matrix metalloproteinases and urokinase-type plasminogen activator-mediated extracellular matrix degradation in human osteosarcoma cell line MG63 by interleukin-1 alpha. J Bone Miner Res. 1995;10: 1374-1384.ArticlePubMedPDF
- 26. Panagakos FS, Kumar S. Modulation of proteases by extracellular matrix and tumor necrosis factor-alpha. Inflammation. 1995;19: 423-443.ArticlePubMedPDF
- 27. Lin SK, Wang CC, Huang S, Lee JJ, Chiang CP, Lan WH, Hong CY. Induction of dental pulp fibroblast matrix metalloproteinase-1 and tissue inhibitor of metalloproteinase-1 gene expression by interleukin-1α and tumor necrosis factor-α through a prostaglandin-dependent pathway. J Endod. 2001;27: 185-189.ArticlePubMed
- 28. Nakata K, Yamasaki M, Iwata T, Suzuki K, Nakane A, Nakamura H. Anaerobic bacterial extracts influence production of matrix metalloproteinases and their inhibitors by human dental pulp cells. J Endod. 2000;26: 410-413.ArticlePubMed
- 29. Shin SJ, Lee JI, Baek SH, Lim SS. Tissue levels of matrix metalloproteinases in pulps and periapical lesions. J Endod. 2002;28: 313-315.ArticlePubMed
- 30. Douglas DA, Shi YE, Sang QA. Computational sequence analysis of the tissue inhibitor of metalloproteinase family. J Protein Chem. 1997;16: 237-255.ArticlePubMedPDF
- 31. Dean DD, Martel-Pelletier J, Pelletier JP, Howell DS, Woessner JF Jr. Evidence for metalloproteinase and metalloproteinase inhibitor imbalance in human osteoarthritic cartilage. J Clin Invest. 1989;84: 678-685.ArticlePubMedPMC
- 32. Johansen JS, Williamson MK, Rice JS, Price PA. Identification of proteins secreted by human osteoblastic cells in culture. J Bone Miner Res. 1992;7: 501-512.ArticlePubMedPDF
- 33. Uchida M, Shima M, Shimoaka T, Fujieda A, Obara K, Suzuki H, Nagai Y, Ikeda T, Yamato H, Kawaguchi H. Regulation of matrix metalloproteinases(MMPs) and tissue inhibitors of metalloproteinases (TIMPs) by bone resorptive factors in osteoblastic cells. J Cell Physiol. 2000;185: 207-214.ArticlePubMed
- 34. Wahl LM, Olsen SE, Sandberg AL, Mergenhagen SE. Prostaglandin regulation of macrophage collagenase production. Proc Natl Acad Sci USA. 1977;74: 4955-4958.ArticlePubMedPMC
- 35. Buck RA, Cai J, Eleazer PD, Staat RH, Hurst HE. Detoxification of endotoxin by endodontic irrigants and calcium hydroxide. J Endod. 2001;27: 325-327.ArticlePubMed
REFERENCES
Figure 1

Amplification plots of real time PCR for MMP-1. Dashed line (--x--) indicates the threshold cycle.
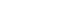
Figure 2
Amplification plots of real time PCR for TIMP-1. Dashed line (--x--) indicates the threshold cycle.
Figure 3
MMP-1 gene expression relative to GAPDH at different LPS concentrations (µg/ml) and different periods of time.
Figure 4

TIMP-1 gene expression relative to GAPDH at different LPS concentrations (µg/ml) and different periods of time.
Figure 5
MMP-1 gene expression relative to GAPDH in osteoblasts stimulated with 10 µg/ml of P. nigrescens LPS pretreated with Ca(OH)2 at different periods of time.
Figure 6
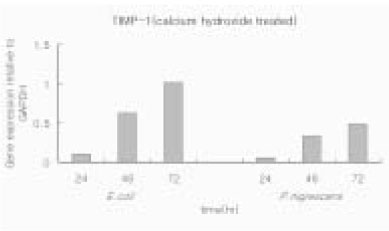
TIMP-1 gene expression relative to GAPDH in osteoblasts stimulated with 10 µg/ml of P. nigrescens LPS pretreated with Ca(OH)2 at different periods of time.
Table 1
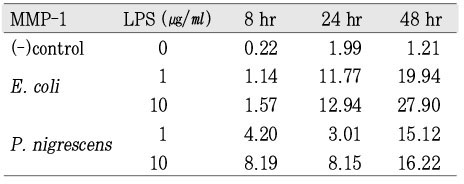
The ratio between the mRNA expression of MMP-1 and GAPDH in osteoblast stimulated with P. nigrescens LPS. The data were acquired in triplicate of each sample.
Table 2
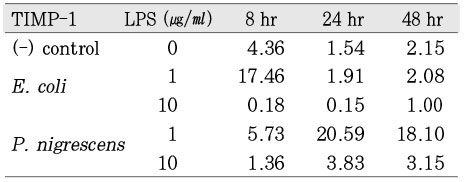
The ratio between the mRNA expression of MMP-1 and GAPDH in osteoblast stimulated with P. nigrescens LPS. The data were acquired in triplicate of each sample.
Tables & Figures
REFERENCES
Citations
Citations to this article as recorded by 
 ePub Link
ePub Link Cite
CiteMMP-1 and TIMP-1 production in MG-63 cells stimulated with Prevotella nigrescens lipopolysaccharide

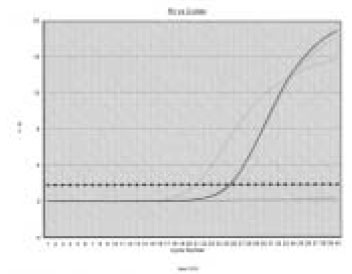
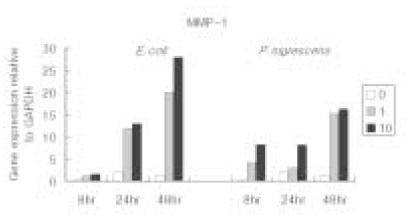
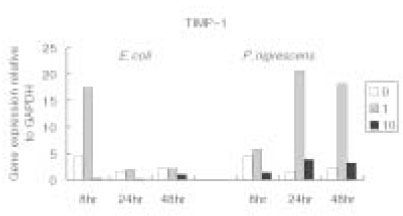
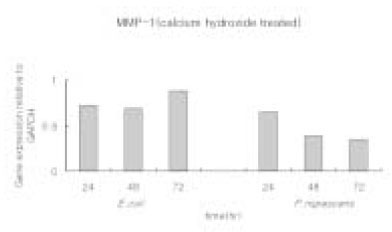
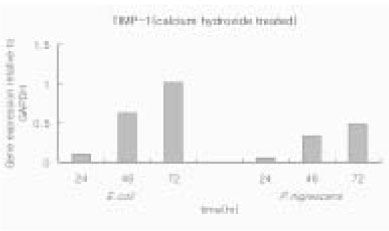
Figure 1
Amplification plots of real time PCR for MMP-1. Dashed line (--x--) indicates the threshold cycle.
Figure 2
Amplification plots of real time PCR for TIMP-1. Dashed line (--x--) indicates the threshold cycle.
Figure 3
MMP-1 gene expression relative to GAPDH at different LPS concentrations (µg/ml) and different periods of time.
Figure 4
TIMP-1 gene expression relative to GAPDH at different LPS concentrations (µg/ml) and different periods of time.
Figure 5
MMP-1 gene expression relative to GAPDH in osteoblasts stimulated with 10 µg/ml of P. nigrescens LPS pretreated with Ca(OH)2 at different periods of time.
Figure 6
TIMP-1 gene expression relative to GAPDH in osteoblasts stimulated with 10 µg/ml of P. nigrescens LPS pretreated with Ca(OH)2 at different periods of time.
Figure 1
Figure 2
Figure 3
Figure 4
Figure 5
Figure 6
MMP-1 and TIMP-1 production in MG-63 cells stimulated with Prevotella nigrescens lipopolysaccharide
The ratio between the mRNA expression of MMP-1 and GAPDH in osteoblast stimulated with P. nigrescens LPS. The data were acquired in triplicate of each sample.
The ratio between the mRNA expression of MMP-1 and GAPDH in osteoblast stimulated with P. nigrescens LPS. The data were acquired in triplicate of each sample.
The ratio between the mRNA expression of MMP-1 and GAPDH in osteoblast-like cells stimulated with P. nigrescens LPS pretreated with Ca(OH)2. The data were acquired in triplicate of each sample.
The ratio between the mRNA expression of TIMP-1 and GAPDH in osteoblast-like cells stimulated with P. nigrescens LPS pretreated with Ca(OH)2. The data are acquired in triplicate of each sample.
Table 1
The ratio between the mRNA expression of MMP-1 and GAPDH in osteoblast stimulated with P. nigrescens LPS. The data were acquired in triplicate of each sample.
Table 2
The ratio between the mRNA expression of MMP-1 and GAPDH in osteoblast stimulated with P. nigrescens LPS. The data were acquired in triplicate of each sample.
Table 3
The ratio between the mRNA expression of MMP-1 and GAPDH in osteoblast-like cells stimulated with P. nigrescens LPS pretreated with Ca(OH)2. The data were acquired in triplicate of each sample.
Table 4
The ratio between the mRNA expression of TIMP-1 and GAPDH in osteoblast-like cells stimulated with P. nigrescens LPS pretreated with Ca(OH)2. The data are acquired in triplicate of each sample.