Articles
- Page Path
- HOME > Restor Dent Endod > Volume 28(5); 2003 > Article
- Original Article The effect of substance P on the secretion of interleukin-8 and MCP (Monocyte Chemoattractant Protein)-1 from the Human Dental Pulp Cells
- Sang-Hyuk Park
-
2003;28(5):-418.
DOI: https://doi.org/10.5395/JKACD.2003.28.5.409
Published online: September 30, 2003
Department of Conservative Dentistry, College of Dentistry, Kyung Hee University, Korea.
- Corresponding author (consdent94@hanmail.net)
Copyright © 2003 Korean Academy of Conservative Dentistry
- 1,481 Views
- 0 Download
I. INTRODUCTION
A common pulpal response to mechanical injury (cavity preparation), bacterial invasion, or bacterial products(dental caries and dental plaque) is characterized by the influx of leukocytes to the affected sites1,2). The consequence of this leukocyte infiltration depends on the degree of the external insults, which may be the function of the tissue, damage the cells, or in the most extreme situation, lead to total necrosis. The types of inflammatory pulpal conditions that are seen in response to external stimuli at cellular levels have been well described in earlier studies3).
Neutrophils are the most predominant cell population that migrates into inflammatory lesions. Besides their protective role, they are involved in tissue damage under pathological conditions because of their high contents in neutral and acid proteases and their ability to generate superoxide and other reactive oxygen derivatives. Circulating neutrophils are quiescent, and most of them are eliminated from the circulation without ever engaging in functional activity. Upon inflammation neutrophils are recruited into affected tissue through a process of multiple activation events that eventually lead to the release of antimicrobial and inflammatory products.
Neutrophil recruitment depends on chemotactic agonists that are synthesized and released at the site of inflammation; e.g., in a periodontal lesion. Chemotactic agonists may be derived from the host or from infecting microorganisms.
The recent discovery of a family of mediators called chemokines, which play a pivotal role in regulating inflammatory reactions, has provided an insight into the molecular mechanisms underlying the kinetics of inflammation. However, the characterization of chemokine expression in the pulp during inflammation and infection has been sparse4).
Interleukin-8(IL-8) was originally described as a 72 amino acid peptide produced by human peripheral blood monocytes stimulated in culture with E.coli lipopolysaccharide5). IL-8 is a member of the chemokine C-X-C subfamily that displays potent chmotactic activities for human neutrophils and T lymphocytes5,6). In addition to chemotaxis, IL-8 induces neutrophil degranulation, resulting in the release of enzymes that cause tissue destruction7).
The activity of IL-8 is not confined to neutrophils. IL-8 stimulates neutrophils to transport CD11/CD18 (LFA-1 or Mac) integrin from the neutrophil cytoplasm to its cell surface8,9). This facilitates neutrophil migration into the tissue. Neutrophil migration may be induced by a haptotactic gradient of IL-8 either on the endothelial cell surface or in the extracellular matrix9). The release of the mature protein and the expression of IL-8 mRNA was demonstrated in endothelial cells, fibroblasts from different tissues, synovial cells, chondrocytes, several types of epithelial cells, and various tumor cells5). IL-8 can be induced by Interleukin-1α(IL-1α), IL-1β, and tumor necrosis factor-α(TNF-α) in many different cells effectively5).
Dental pulp is a richly innervated tissue. The sensory fibers terminate into the predentin layer of the tooth, together with the fluid in the dentinal tubules, rendering the tooth hypersensitive to irritations from either caries or mechanical/chemical treatments during dental procedures.
These biological or iatrogenic insults can cause dental pain which is largely due to the release of mediators in the pulp in response to the irritations. Particularly, pain due to pulpal hyperemia-the first sign of inflammation in pulp, is the result of dilatation of blood vessels leading to increased tissue pressure10).
The richly innervated dental pulps contain nerve fibers expressing substance P(SP) or calcitonin gene-related peptide(CGRP)11). When pulp is directly injured such as pulp exposure, neuropeptide levels increase in the pulp and trigeminal ganglia12,13). The neuropeptides may be released upon stimulation from dental procedures or caries invasion14), causing increased vasodilatation and pulpal blood flow15). Subsequently, influx of inflammatory cells from blood into tissue ensues which process is highly regulated. The local tissue cells and endothelial cells play a pivotal role in this regulation16). Interleukin-8(IL-8), a potent neutrophil chemoattractant, is frequently expressed in endothelial cells of inflamed pulps, while rarely expressed in endothelial cells of normal pulps17). This suggests the key regulatory role of IL-8 expressed by endothelial cells in controlling the influx of leukocytes into the pulp tissue.
SP or CGRP induces human corneal epithelial cells to secrete IL-8, but does not induce to secrete monocyte chemoattractant protein-1(MCP-1) or RANTES18,19). SP or CGRP induces the synthesis of IL-6, IL-8 and TNF-α in bronchial epithelial cells20), and upregulates IL-8 RA mRNA in dermal fibroblasts and keratinocytes21). SP induces IL-8 production by osteoarthritis fibroblasts, while CGRP increases IL-8 and IL-6 secretion from rheumatoid arthritis fibroblasts22).
Neurogenic inflammation has been recognized to play an important role in initiating and sustaining of pulp inflammation23). The pulpal innervation may modulate several aspects of the inflammatory response via secretion of neuropeptides. In this present study, these neuropeptides that may be questioned about roles in recruiting leukocytes by inducing the release of the chemokine IL-8 in the pulp during inflammation were tested. The response of human pulp cells in releasing IL-8 after the stimulation with SP and/or CGRP were investigated.
II. MATERIALS AND METHODS
Freshly extracted, intact, caries-free third molars were obtained from the patients(15~25 years old) in the Department of Oral Surgery at the UCLA School of Dentistry according to a protocol approved by the UCLA Medical Institutional Review Board. Immediately after extraction, teeth were stored in phosphate buffered saline(PBS) and transferred to the laboratory. Under water coolant, each tooth was grooved longitudinally with a fissure bur at high speed. The tooth was then split with the driver, and the entire pulp(coronal and radicular) elevated with cotton pliers, maximum pulpal tissue was obtained. Some samples were divided into several small fragments approximately 2×2 mm in size each.
Repeated washing with PBS, pulp fragments were placed in a 60 mm culture dish containing Dulbecco's Modified Eagle Medium(DMEM; Life Technologies/GIBCO BRL, Gaithersburg, MD) supplemented with 10% fetal bovine serum(FBS). Pulp cells from pulp fragments were grown to confluence and passed at 1:2 ratio until used for experiments(passages 3-8 were used). ECV 304 cells(a human endothelial cell line obtained from Dr. H. Shau, UCLA, Los Angeles, CA) were grown in RPMI-1640(Life Technologies/GIBCO BRL, Gaithersburg, MD) with 10% FBS. Cell culture media were supplemented with 100 units/ml penicillin-G, 100 µg/ml streptomycin, and 0.25 µg/ml fungizone (Gemini Bio-Products, Inc., Woodland, CA).
Some samples were divided in half, the 2 halves being stored separately under different conditions according to the analysis used(ELISA or immunocytochemistry). Samples for the ELISA were placed in a microcentrifuge tube containing 0.2 ml of phosphate-buffered saline solution(PBS).
Synthetic human SP, Spantide(SP receptor antagonist) I and α-CGRP(Sigma, St. Louis, MO) were prepared in sterile water with 0.1% low-endotoxin bovine serum albumin(BSA, Sigma). Cells were seeded in 48- or 96-well plates and grown to confluence before used for experiment. Alpha minimal essential medium containing L-glutamine(Life Technologies/GIBCO BRL, Gaithersburg, MD) with 1% FBS was used 24 h before cells were stimulated with neuropeptiedes.
This low serum medium was used to decrease the base-line level of IL-8 secretion from these cells. Based on the preliminary experiments, the serum induced significant levels of IL-8 and MCP-1 from pulp and ECV 304 cells which interfered with the induction effect of SP. Varying doses of the neuropeptides or 0.1% BSA(Mock stimulation) were added to the cell cultures. Recombinant human (rh) TNF-α(R&D Systems, Minneaplis, MN), a known IL-8 and MCP-1 agonist, was used as a positive control for IL-8 and MCP-1 induction.
To determine the kinetics of IL-8 and MCP-1 induction by SP, 24 h studies in the time course were performed in which the supernatant was collected and fresh medium added at every 4 h interval. IL-8 induction by either only the first 4 h of SP stimulation or continuous presence of SP throughout the 24 h study period were tested. In order to detect increased IL-8 at each time-point(4 h interval), a high concentration of SP(10-4 M) was used.
As previously described24), optimal concentrations of polyclonal goat anti-human IL-8 antibodies (R&D Systems, Minneaplis, MN) and monoclonal anti-human MCP-1 antibodies(R&D System, Minneapolis, MN) were used as capturing antibodies, polyclonal rabbit anti-human IL-8 antibodies (Endogen Inc., Cambridge, MA) and polyclonal rabbit anti-human MCP-1 antibodies(Cell Science, Inc. Norwood, MA) as detecting antibodies, and horseradish peroxidase (HRPO)-labeled polyclonal goat anti-rabbit immunoglobulin G (Biosource International, Camarillo, CA) as a second-step antibody. Bound HRPO was visualized with fresh developing buffer containing substrate of optimal concentrations of 3, 3', 5, 5'-Tetramethylbenzidine (Sigma), H2O2 and sodium acetate, pH 6.0. The developing reaction was stopped with the addition of 1.2 M sulfuric acid. Absorbance was determined at 450 nm with a microplate reader(Bio-Tek Instrument, Inc., Laguna Hills, CA) and concentrations were determined by the Delta Soft III software(Bio-Tek Instrument, Inc.). Purified rh IL-8(Endogen Inc., Cambridge, MA) and purified rh MCP-1(R&D System, Minneapolis, MN) of known concentration was used to establish a standard curve for determining concentrations of the experimental samples.
III. RESULTS
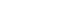
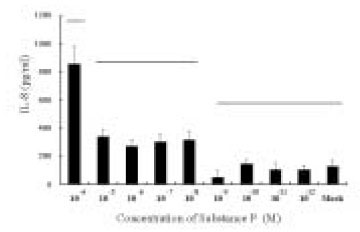
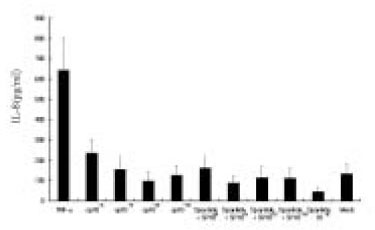
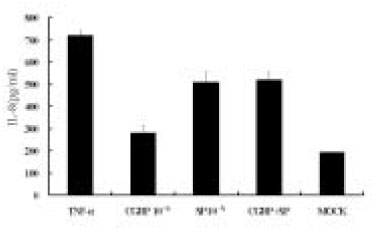
The dose response studies presented in Fig. 1 show that, comparing to Mock stimulation, IL-8 secretion started to increase when SP concentration was higher than 10-12 M. There was 3.8-fold increase at 10-8 M of SP stimulation(t test, p<0.05) and more increase(13.8-fold) was observed when 10-4 M of SP was used. However, no or minimal increase of IL-8 production with CGRP(up to 10-4 M) stimulation was observed(statistically insignificant, data not shown). To determine the specificity of this IL-8 induction by SP, an antagonist(Spantide I) to SP receptor NK-1 was utilized. In the presence of Spantide(10-8 to 10-6 M), IL-8 induction by SP(10-5 M) was inhibited, indicating the specificity of this induction(Fig. 2). This also suggests that the main SP receptor on the cultured pulp cells appears to be NK-1.
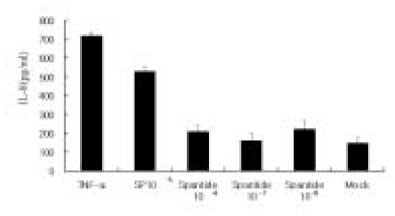
Since endothelial cells play a major role in regulating local inflammation, whether SP can induce endothelial cells to secrete IL-8 was tested. The isolation of human pulpal endothelial cells has not been reported. Therefore, an endothelial cell line from the human umbilical vein was used for the study. As shown in Fig. 3, IL-8 was only mildly induced in ECV304 cells at a high concentration of SP(10-4 M). The NK-1 receptor antagonist also inhibited SP stimulation of IL-8 in these cells as in pulp cells.
Although CGRP alone did not significantly induce IL-8, it may synergistically enhance the IL-8 induction capacity of SP. To test this possibility, high concentrations of both neuropeptides(10-5 M) were used.
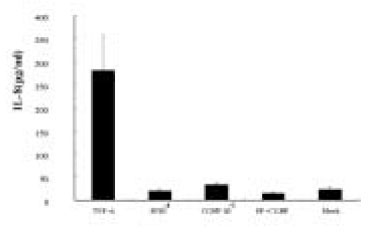
The results shown in Fig. 4 and Fig. 5 suggest that SP plus CGRP did not further induce, but appear to reduce IL-8 secretion from pulp cells or ECV 304 cells compared with SP alone. This indicates that there is no synergistic effect on IL-8 induction by SP plus CGRP.
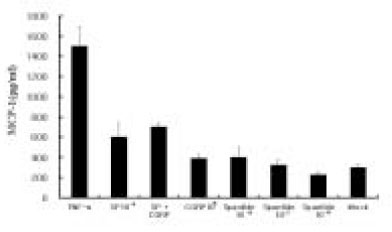
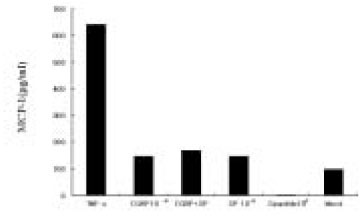
After IL-8 ELISA, the supernatant was stored at -80℃ for MCP-1 ELISA. Dose experimental studies showed MCP-1 secretion was 2-fold higher at 10-4 M of SP stimulation(One-way ANOVA test, p<0.05) comparing to Mock stimulation(Data not shown). Fig. 6 shows that MCP-1 was only mildly induced in pulp cells at 10-5 M of SP, CGRP alone did not significantly induce MCP-1, and CGRP plus SP induction was not significantly higher than Mock stimulation. At the concentration of Spantide(10-8 M), the NK-1 receptor antagonist also inhibited SP(10-5 M) stimulation of MCP-1 in the pulp cells. As shown in Fig. 7, MCP-1 was only mildly induced in ECV 304 cells at a high concentration of SP(10-5 M), CGRP(10-5 M) compared with Mock, and SP plus CGRP(both 10-5 M) in ECV 304 cells mildly induced MCP-1 compared with SP(10-5 M) or CGRP(10-5 M). This indicates that there is no significant synergistic effect on MCP-1 induction by SP plus CGRP. In the presence of Spantide(10-7 M), MCP-1 induction by SP (10-5 M) was inhibited (Fig. 7).
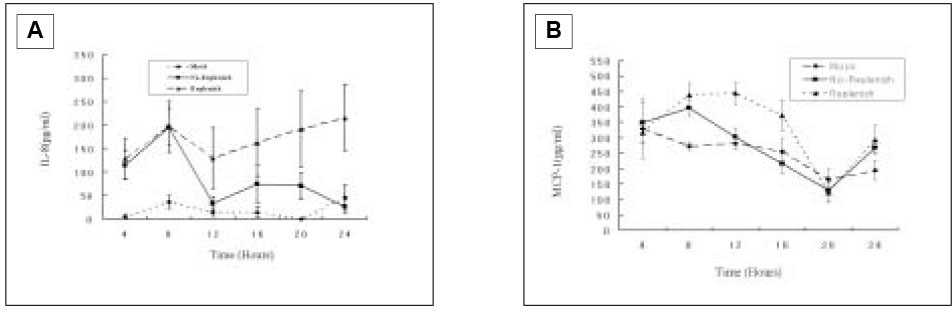
When pulp cells were stimulated with SP during the first 4 h, IL-8 secretion increased after 4 h and reached the maximal level at 8 h, followed by decreasing down to only slightly higher than the base-line level after 12 h. When SP was present during the entire course, i.e., fresh medium containing SP was added at every time point after collection of the supernatant, IL-8 secretion reached the maximal level at 8 h and appeared to remain at the level during the subsequent periods of the study(Fig. 8A).
After IL-8 ELISA test, the supernatant was kept at -80℃ for the further MCP-1 ELISA. As shown in Fig. 8B, there was no MCP-1 induction from the pulp cells with SP(10-4 M) after the first 4 h and reached the maximal level at 8 h, followed by decreasing down to only slightly higher than the base-line level after 12 h. When SP was present during the entire course, MCP-1 secretion reached the maximal level at 8 to 12 h, and decreased down to only slightly higher than the base-line level after 16 h.
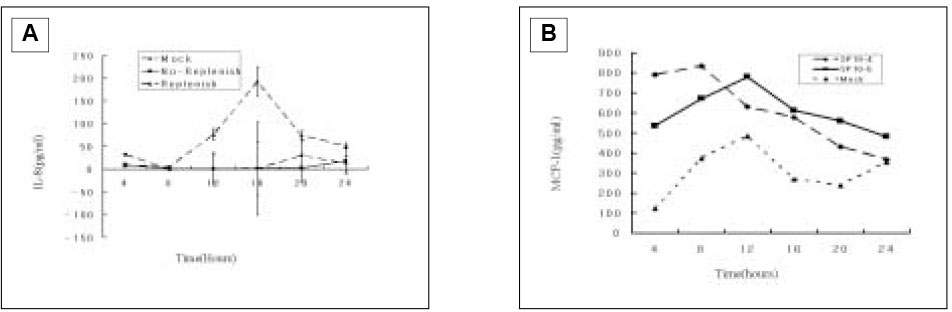
The studies in the course of time for ECV304 cells showed that no or minimal induction was detected throughout the 24 h period when SP was only present in the first 4 h of incubation(Fig. 9A). This was not surprising since IL-8 induction was very weak based on the studies shown in Fig. 3 in which there was a 12 h accumulation of IL-8. Therefore, a 4 h interval would have little accumulation of IL-8 to detect. Nonetheless, when SP was continuously present in the cell culture, moderate IL-8 induction was detected.
Because studies in the course of time for ECV 304 cells showed minimal IL-8 induction, in order to detect increased MCP-1, only two different concentrations of SP(10-4 and 10-5 M) were used by first 4 h stimulation. In the result of Fig. 9B, 10-4 M of SP increased MCP-1 secretion than 10-5 M of SP within 4 h, and 10-5 M of SP increased up MCP-1 secretion higher than 10-5 M of SP by 8 h showing maximal level of MCP-1 secretion at 12 h, followed by decreasing down to the base-line level at 24 h.
IV. DISCUSSION
These demonstrated data that human pulp cells grown out of the dental pulp fragments increased the secretion of IL-8 in response to SP stimulation suggest a more direct role of SP in initiation of inflammatory cell infiltration in dental pulp. The failure of CGRP to induce significant IL-8 correlates with the concept that generally SP upregulates immune and inflammatory response, whereas CGRP inhibits these responses25). It may be related with the results that pulpal levels of CGRP were higher than SP25,32). Apparently, CGRP does not participate in the initiation of inflammatory infiltration in the pulp. In addition, SP plus CGRP seem to have no synergistic effect on IL-8 induction in pulp cells.
While the concentrations of SP needed to obtain statistically significant IL-8 induction in pulp cells under the experimental settings were between 10-8 and 10-4 M, induction was observed at lower concentration of SP(10-12 M) stimulation. This suggests that in the inflamed pulp tissue where SP concentration reaches approximately 10-8 to 10-9 M (Hargreaves personal communication, 2002)26), local pulp cells may be induced to secrete IL-8 by the SP. In many cell system, ranges of 10-6 to 10-9 M of SP concentration are needed to induce significant cytokines production including IL-8 in vitro18-21). Based on these data, SP at 10-8 M induced IL-8 approximately 4-fold, suggesting that SP is a moderate IL-8 stimulator for pulp cells. The endothelial cells(ECV 304) utilized in these studies showed only mild potential to increase the production of IL-8 upon high concentration of SP(10-4 M) stimulation. If pulpal endothelial cells behave the same way as ECV 304 cells in terms of IL-8 response with the stimulation of SP, it is unlikely that these cells will produce this chemokine when SP is released in the pulp. It remains to be shown whether pulpal endothelial cells in vivo are sensitive to SP stimulation of IL-8 production. It should be noted that IL-8 secretion from pulp cells may be internalized by the nearby endothelial cells and eventually be presented on the luminal side of the endothelial cells and interact with its receptor on leukocytes in the blood stream27-29).
Nerve fibers in the pulp associated with large blood vessels, subodontoblast layer, between odontoblasts, or the predentin and dentin contain SP11). Dental procedures which stimulate sensory response may cause the local release of SP and other mediators in the pulp. Based on these present findings, the localized pulp cells adjacent to these sensory nerve fibers may express IL-8 in a timely manner in response to the released SP from the nerve fibers. These studies in the course of time show that IL-8 induction in pulp cells occurs rapidly within 4 h upon SP stimulation in consistent with the general feature of IL-8 induction kinetics by other stimuli30). Monocyte chemoattractant protein-1(MCP-1) was also tested to evaluate whether it may be induced in pulp cells by SP and/or CGRP.
The results indicated that neither SP nor CGRP induced MCP-1 significantly at physiological concentration(10-8 to 10-9 M). Only mild to moderate MCP-1 induction was observed at 10-4 M for both neuropeptides.
SP and CGRP increased IL-1β, IL-6, and TNF-α from cultured lymphocyte in a concentration-dependent manner, and synergistically increased basal TNF-α secretion at 10-7 M of SP and CGRP33). In this study, there were no synergic effect for SP and CGRP(10-5 M) to secrete IL-8 and MCP-1 on the pulp cells.
Substance P receptor antagonist, Spantide(10-6 M) decreased proline level compared with SP(10-8 M) in rat bone marrow-derived osteogenic cells, but there was no statistically significant difference31). Spantide(10-7 M) blocked the secretion of IL-8 with the stimulation of SP(10-5 M) from the pulp cells, and also inhibited the secretion of MCP-1 at 10-8 M of Spantide.
Other proinflammatory cytokines produced during inflammation also regulate the production of neuropeptides in a positive feedback loop. SP increases the secretion of IL-1, TNF-α, and IL-6 from macrophages and mast cells34). CGRP plays an important role in the healing response of pulpal tissue after injury35). Thus, upon the release of other proinflammatory cytokines, SP may be produced by pulp cell persistently for an extensive amount of time which may generate significant clinical symptoms. In conclusion, the present study indicates that SP induced the secretion of IL-8 and MCP-1 in the dental pulp cells and endothelial cells(ECV 304 cells), and the secretion of neuropeptide was highest at 8 to 12 hr after the stimulation of SP in the course of time. However, CGRP did not induce the IL-8 and MCP-1 in the dental pulp cells.
V. CONCLUSION
The purpose of the present study was to examine the coordinate activity between neuropeptide and cytokine, and their important role in sensing and eliciting rapid immune response to the external irritations to the dental pulp. For the purpose, the induction of the IL-8 and MCP-1 by the stimulation of neuropeptide in the pulp cells and endothelial cells(ECV 304 cells) was done in the dose dependent manner and in the course of time. The specificity in the dental pulp cells and endothelial cells(ECV304 cells) was measured using Spantide (SP antagonist).
According to this study, the results were as follows:
When SP concentration was higher than 10-8 M, IL-8 secretion from human dental pulp cells was significantly increased comparing to mock stimulation(p<0.05). And IL-8 secretion from human dental pulp cells at 10-4 M of SP was significantly higher than lower concentration of SP(10-5 to 10-8 M)(p<0.05).
SP(10-5 M) stimulation of the human dental pulp cells to secrete IL-8 and MCP-1 was inhibited by Spantide(10-5 M).
When the human dental pulp cells were stimulated by CGRP(10-4 M), IL-8 secretion was not increased, and synergistic induction of IL-8 with SP(10-5 M) plus CGRP(10-5 M). There was no synergistic induction of MCP-1 from human dental pulp cells with SP(10-5 M) plus CGRP(10-5 M) comparing with SP(10-5 M) only.
There was only mild induction of IL-8 and MCP-1 in the endothelial cells(ECV 304 cells).
SP stimulation of endothelial cells(ECV 304 cells) to secrete IL-8 and MCP-1 was inhibited by Spantide.
Pulp cells were stimulated with SP(10-4 M) at every 4 hours during 24 hours in the course of time, and IL-8 secretion was measured, which increased at 4 hours and reached the maximal level at 8 hours(p<0.05), MCP-1 secretion was maximum at 8 to 12 hours of SP stimulation(10-4 M).
These results suggest that SP played an important role of IL-8 and MCP-1 induction from human dental pulp cells, but CGRP did not induce any secretion. And SP may have positive relation with the inflammation of the human dental pulp cells.
- 1. Stanley HR. Importance of the leukocyte to dental health. J Endod. 1977;3: 334-341.PubMed
- 2. Bergenholtz G, Lindhe J. Effect of soluble plaque factors on inflammatory reactions in the dental pulp. Scand J Dent Res. 1975;83: 153-158.ArticlePubMed
- 3. Seltzer S, Bender IB, Ziontz M. The dynamics of pulp inflammation : Correlations between diagnostic data and actual histologic findings in the pulp. Oral Surg Oral Med Oral Pathol. 1963;16: 846-871.ArticlePubMed
- 4. Rauschenberger C, Bailey J, Cootauco C. Detection of human IL-2 in normal and inflamed dental pulps. J Endod. 1997;23: 366-370.ArticlePubMed
- 5. Baggiolini M, Walz A, Kunkel SL. Neutrophil-activating peptide-1/interleukin 8, a novel cytokine that activates neutrophils. J Clin Invest. 1989;84: 1045-1049.ArticlePubMedPMC
- 6. Larsen CG, Anderson AO, Appella E, Oppenheim JJ, Matsushima K. The neutrophil-activating protein(NAP-1) is also chemotactic for T lymphocytes. Science. 1989;243: 1464-1466.ArticlePubMed
- 7. Peveri P, Walz A, Dewald B, Baggiolini M. A novel neutrophil activating factor produced by human mononuclear phagocytes. J Exp Med. 1988;167: 1547-1559.ArticlePubMedPMCPDF
- 8. Detmers PA, Lo SK, Olsen-Egbert E, Walz A, Baggiolini M, Cohn ZA. Neutrophil-activating protein 1/interleukin 8 stimulates the binding activity of the leukocyte adhesion receptor CD11b/CD18 on human neutrophils. J Exp Med. 1990;171: 1155-1162.ArticlePubMedPMCPDF
- 9. Rot A. Neutrophol attractant/activation protein-1 (interleukin-8) induces in vito neutrophil migration by haptotactic mechanism. Eur J Immunol. 1993;23: 303-306.ArticlePubMed
- 10. Kim S. Neurovascular interactions in the dental pulp in health and inflammation. J Endod. 1990;16: 48-53.ArticlePubMed
- 11. Wakisaka S. Neuropeptides in the dental pulp : distribution, origins, and correlation. J Endod. 1990;16: 67-69.ArticlePubMed
- 12. Grutzner EH, Garry MG, Hargreaves KM. Effect of injury on pulpal levels of immunoreactive substance P and immunoreactive calcitonin gene-related peptide. J Endod. 1992;18: 553-557.ArticlePubMed
- 13. Buck S, Reese K, Hargreaves KM. Pulpal exposure alters neuropeptide levels in inflamed dental pulp and trigeminel ganglia: evaluation of axonal transport. J Endod. 1999;25: 718-721.PubMed
- 14. Rodd HD, Boissonade FM. Comparative immunohistochemical analysis of the peptidergic innervation of human primary and permanent tooth pulp. Arch Oral Biol. 2002;47: 375-385.ArticlePubMed
- 15. Olgart LM, Edwall B, Gazelius B. Neurogenic mediators in control of pulpal blood flow. J Endod. 1989;15: 409-412.ArticlePubMed
- 16. Muller WA, Randolph GJ. Migration of leukocytes across endothelium and beyond : molecules involved in the transmigration and fate of monocytes. J Leukoc Biol. 1999;66: 698-704.ArticlePubMedPDF
- 17. Huang GTJ, Chugal N, Potente AP, Zhang X. Constitutive Expression of Interleukin-8 and Intercellular Adhesion Molecule-1 in Human Dental Pulps. Int J Oral Biol. 1999;24: 163-168.
- 18. Tran MT, Ritchie MH, Lausch RN, Oakes JE. Calcitonin gene-related peptide induces IL-8 synthesis in human corneal epithelial cells. J Immunol. 2000;164: 4307-4312.ArticlePubMedPDF
- 19. Tran MT, Lausch RN, Oakes JE. Substance P differentiall stimulates IL-8 synthesis in human corneal epithelialcells. Invest Ophthalmol Vis Sci. 2000;41: 3871-3877.PubMed
- 20. Veronesi B, Carter JD, Devlin RB, Simon SA, Oortgiesen M. Neuropeptides and capsaicin stimulate the release of inlfammatory cytokines in a human broncheal epithelial cell line. Neuropeptides. 1999;33: 447-456.ArticlePubMed
- 21. Kiss M, Kemény L, Gyulai R, Michel G, Husz S, Kovács R, Dobozy A, Ruzicka T. Effects of the neuropeptides substance P, calcitonin gene-related peptide and alpha-melanocyte-stimulating hormone on the IL-8/IL-receptor system in a cultured human keratinocyte cell line and dermal fibroblasts. Inflammation. 1999;23: 557-567.ArticlePubMedPDF
- 22. Raap T, Justen HP, Miller LE, Cutolo M, Scholmerich J, Straub RH. Neurotransmitter modulation of interleukin 6 (IL-6) and IL-8 secretion of synovial fibroblasts in patients with rheumatoid arthritis compared to osteoarthritis. J Rheumatol. 2000;27: 2558-2565.PubMed
- 23. Byers MR, Närhi MV. Dental injury models; experimental tools for understanding neuroinflammatory interactions and polymodal nociceptor functions. Crit Rev Oral Biol Med. 1999;10: 4-39.ArticlePubMedPDF
- 24. Huang GT, Potente AP, Kim JW, Chugal N, Zhang X. Increased interleukin-8 espression in inflamed human dental pulps. Oral Surg Oral Med Oral Pathol Oral Radiol Endod. 1999;88: 214-220.PubMed
- 25. Stashenko P, Teles R, D'Souza R. Periapical inflammatory responses and their modulation. Crit Rev Oral Biol Med. 1998;9: 498-521.ArticlePubMedPDF
- 26. Awawdeh L, Lundy FT, Shaw C, Lamey PJ, Linden GJ, Kennedy JG. Quantitative analysis of substance P, neurokinin A and calcitonin gene-related peptide in pulp tissue from painful and healthy human teeth. Int Endod J. 2002;35: 30-36.ArticlePubMed
- 27. Rot A, Hub E, Middleton J, Pons F, Rabeck C, Thierer K, Wintle J, Wolff B, Zsak M, Dukor P. Some aspects of IL-8 pathophysiology. III: Chemokine interaction with endothelial cells. J Leukocyte Biol. 1996;59: 39-44.ArticlePubMedPDF
- 28. Middleton J, Neil S, Wintle J, Clark-Lewis I, Moore H, Lam C, Auer M, Hub E, Rot A. Transcytosis and surface presentaion of IL-8 by venular endothelial cells. Cell. 1997;91: 385-395.ArticlePubMed
- 29. Wolff B, Burns AR, Middleton J, Rot A. Endothelial cell "memory" of inflammatory stimulation: human venular endothelial cells store interleukin 8 in Weibel-Palade bodies. J Exp Med. 1998;188: 1757-1762.ArticlePubMedPMCPDF
- 30. Huang GT, Kim D, Lee JK, Juramitsu HK, Haake SK. Interleukin-8 and intercellular adhesion molecule 1 regulation in oral epithelial cells by selected periodontal bacteria: multiple effects of Porphyromonas gingivalis via antagonistic mechanisms. Infect Immun. 2001;69: 1364-1372.ArticlePubMedPMCPDF
- 31. Adamus MA, Dabrowski ZJ. Effect of the Neuropeptide Substance P on the rat bone marrow-derived osteogenic cells in vitro. J Cell Biochem. 2001;81: 499-506.ArticlePubMed
- 32. Hargreaves KM, Walter RB, Garry MG. An in vitro method to evaluated regulation of neuropeptide release from dental pulp. J Endod. 1992;18: 597-600.PubMed
- 33. Cuesta MC, Quintero H, Pons H, Suarez-Roca . Substance P and calcitonin gene-related peptide increase IL-1β, IL-6 and TNF-α secretion from human peripheral blood mononuclear cells. Neurochem Int. 2002;40: 301-306.ArticlePubMed
- 34. Ansel JC, Armstrong CA, Song I, Quinlan KL, Olerud JE, Caughman SW, Bunnett NW. Interaction of the skin and nervous system. J Investig Dermatol Symp Proc. 1997;2: 23-26.PubMed
- 35. Taylor PE, Byers MR. An immunocytochemical study of the morphological reaction of nerves containing calcitonin gene-related peptide to microabscess formation and healing in rat molars. Arch Oral Biol. 1990;35: 629-638.ArticlePubMed
REFERENCES
Tables & Figures
REFERENCES
Citations
Citations to this article as recorded by 
 ePub Link
ePub Link Cite
CiteThe effect of substance P on the secretion of interleukin-8 and MCP (Monocyte Chemoattractant Protein)-1 from the Human Dental Pulp Cells
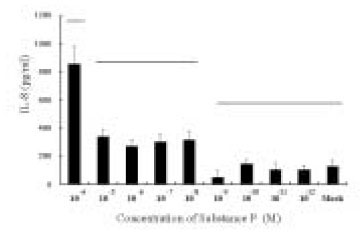
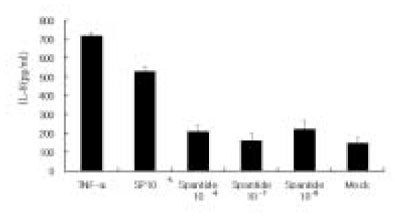
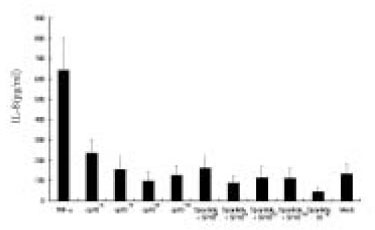
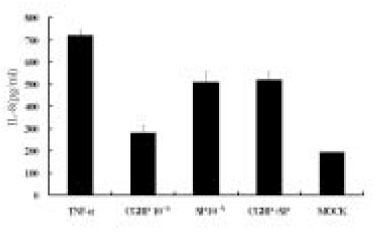
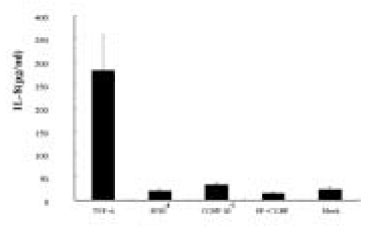
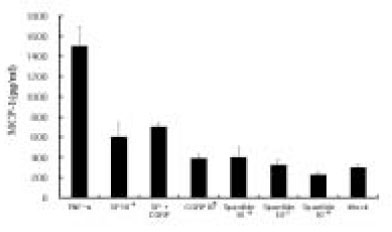
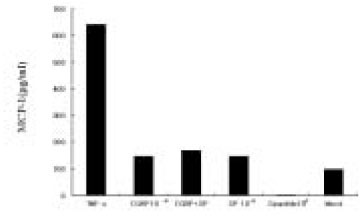
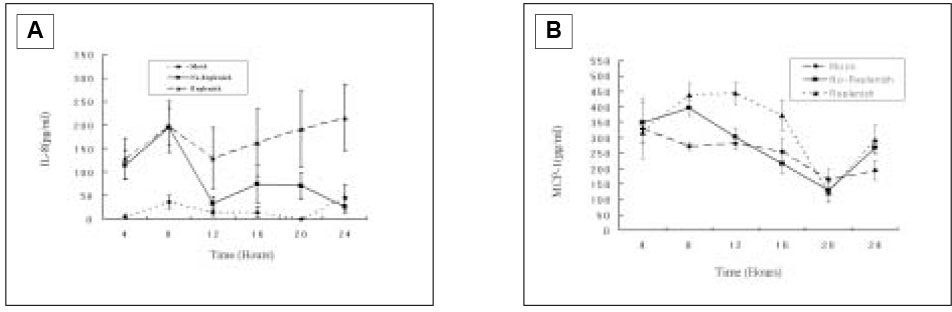
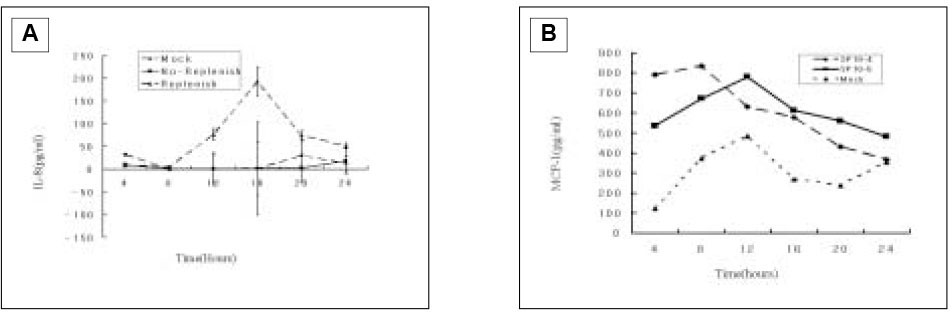
Fig. 1
Dose response of IL-8 secretion in human pulp cells stimulated with sp.
Fig. 2
IL-8 secretion from the pulp cells following SP stimulation.
Fig. 3
IL-8 secretion from ECV 304 cells in response to SP stimulation.
Fig. 4
ELISA detection of IL-8 from the pulp cells stimulated with SP, CGRP, or SP+CGRP.
Fig. 5
ELISA detection of IL-8 from ECV 304 cells stimulated with SP, CGRP, or SP+CGRP.
Fig. 6
MCP-1 secretion from the pulp cells in response to SP stimulation.
Fig. 7
MCP-1 secretion from ECV304 cells in response to SP stimulation.
Fig. 8
IL-8(A) and MCP-1(B) secretion from pulp cells following SP(10-4 M) stimulation in the course of time.
Fig. 9
IL-8 secretion from ECV 304 cells following SP(10-5 M) stimulation(A) and MCP-1 secretion from ECV 304 cells following SP(10-4 and 10-5 M) stimulation(B) in the course of time.
Fig. 1
Fig. 2
Fig. 3
Fig. 4
Fig. 5
Fig. 6
Fig. 7
Fig. 8
Fig. 9
The effect of substance P on the secretion of interleukin-8 and MCP (Monocyte Chemoattractant Protein)-1 from the Human Dental Pulp Cells