Articles
- Page Path
- HOME > Restor Dent Endod > Volume 40(4); 2015 > Article
-
Research Article
Identification of
Enterococcus faecalis antigens specifically expressedin vivo - Seok-Woo Lee1,2, Uttom K. Shet3, Sang-Won Park4, Hyun-Pil Lim4, Kwi-Dug Yun4, Seong Soo Kang5, Se Eun Kim5
-
2015;40(4):-311.
DOI: https://doi.org/10.5395/rde.2015.40.4.306
Published online: October 5, 2015
1Department of Dental Education, Dental Science Research Institute and BK21 Project, School of Dentistry, Gwangju, Korea.
2Department of Periodontology, Dental Science Research Institute and BK21 Project, School of Dentistry, Gwangju, Korea.
3Department of Maxillofacial Surgery, Dental Science Research Institute and BK21 Project, School of Dentistry, Gwangju, Korea.
4Department of Prosthodontics, Dental Science Research Institute and BK21 Project, School of Dentistry, Gwangju, Korea.
5Department of Veterinary Medicine, Chonnam National University, Gwangju, Korea.
- Correspondence to Seok-Woo Lee, DDS, MS, PhD. Professor, Departments of Dental Education and Department of Periodontology, School of Dentistry, Chonnam National University, 77 Yongbong-ro, Buk-gu, Gwangju, Korea 61186. TEL, +82-62-530-5820; FAX, +82-62-530-5810; swlee@jnu.ac.kr
• Received: June 10, 2015 • Accepted: August 10, 2015
©Copyrights 2015. The Korean Academy of Conservative Dentistry.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
- 2,307 Views
- 5 Download
- 4 Crossref
Abstract
-
Objectives Molecular mechanism of the pathogenicity of Enterococcus faecalis (E. faecalis), a suspected endodontic pathogen, has not yet been adequately elucidated due to limited information on its virulence factors. Here we report the identification of in vivo expressed antigens of E. faecalis by using a novel immunoscreening technique called change-mediated antigen technology (CMAT) and an experimental animal model of endodontic infection.
-
Materials and Methods Among 4,500 E. coli recombinant clones screened, 19 positive clones reacted reproducibly with hyperimmune sera obtained from rabbits immunized with E. faecalis cells isolated from an experimental endodontic infection. DNA sequences from 16 of these in vivo-induced (IVI) genes were determined.
-
Results Identified protein antigens of E. faecalis included enzymes involved in housekeeping functions, copper resistance protein, putative outer membrane proteins, and proteins of unknown function.
-
Conclusions In vivo expressed antigens of E. faecalis could be identified by using a novel immune-screening technique CMAT and an experimental animal model of endodontic infection. Detailed analysis of these IVI genes will lead to a better understanding of the molecular mechanisms involved in the endodontic infection of E. faecalis.
Introduction
Bacterial pathogens are known to possess sophisticated mechanisms for their survival within their hosts by expressing specific products. These products, called virulence factors, are involved in the steps of infectious process, resulting in host-pathogen interaction.1,2 The outcome of this complex and dynamic interaction is manifested as signs and symptoms of clinical infectious diseases. In cases of root canal and periapical infections, it is widely accepted that anaerobic Gram-negative bacteria are responsible.3,4,5 However, the specific roles of putative bacterial pathogens in the pathogenesis of endodontic infections have not yet been adequately assessed. Of these putative pathogens, Enterococcus faecalis (E. faecalis) has been implicated to play an important role in persistent root canal and periapical infections mainly due to its unique presence in these lesions.6,7
At present, molecular pathogenesis of E. faecalis infection is not well understood, since the virulence factors involved in root canal and periapical infections have not yet been adequately identified or characterized. Several putative virulence factors of E. faecalis with potential of pathogenicity have been described, including aggregation substance, several surface adhesions, sex pheromones, lipoteichoic acid, gelatinase, hyaluronidase, extracellular superoxide production, and bacteriocins.8 However, their exact roles in endodontic infections have not yet been determined since it is unclear whether these virulence factors are involved in the actual disease process. Additional virulence factors from E. faecalis need be identified and characterized to better understand the interactions between this microorganism and the host since they mediate the infectious activities in root canal and periapical infection.
Most traditional bacterial virulence factors have been identified using an in vitro approach in the laboratory. It is well established that these virulence factors may not be involved in the actual disease process since they are not even expressed inside the host.9 It is therefore crucial to identify and/or characterize genes (proteins) expressed in vivo instead of using an in vitro approach, since these in vivo expressed genes (proteins) are more likely to serve as virulence factors. Numerous molecular techniques have been developed for identifying bacterial genes and their products expressed only in the host, including in vivo expression technology, signature-tagged mutagenesis, differential fluorescence induction, transcriptional and proteomic profiling, and in vivo-induced antigen technology (IVIAT).10,11,12,13,14,15
Change-mediated antigen technology (CMAT), a modification of IVIAT, is an antibody-based screening technique used to identify bacterial antigens expressed only in vivo when bacteria undergo changes such as an infection.16 This technology uses hyperimmune antisera raised against bacterial cells isolated from infected tissues. The hyperimmune antisera are subsequently adsorbed with cells and extracts of cells that grow in vitro to produce a probe that is reactive only with proteins expressed exclusively in vivo. CMAT has not been used to identify antigens of E. faecalis expressed in vivo. Therefore, the objective of this study was to use CMAT to screen and identify in vivo-expressed antigens of E. faecalis using an experimental animal infection model.
Materials and Methods
E. faecalis ATCC 29212 was purchased from American Type Culture Collection (Manassas, VA, USA) and grown on Brain heart infusion (BHI) agar (Becton Dickson, Sparks, MD, USA) at 37℃. For broth growth, bacterial cells were incubated in 10 mL of BHI broth for 12 hours at 37℃. Cell pellets were collected and kept at -80℃ until used. E. coli BL21 (DE3)/pLysS competent cells (Stratagene, La Jolla, CA, USA) were grown overnight on lysogeny broth (LB) agar plates or in LB broth containing kanamycin (30 µg/mL) at 37℃.
Genomic DNA from E. faecalis cells were extracted using a G-spin Genomic DNA Extraction Kit for Bacteria (iNtRON Biotechnology, Sungnam, Korea) according to manufacturer's instructions. Purified genomic DNA was then randomly sheared by sonication using a SONOPULS Ultrasonic homogenizer (Bandelin, Berlin, Germany) for 6 seconds at 100% power set of Sonic to generate DNA fragments sized of 1 - 5 kb. After fractionated by agarose gel electrophoresis to remove fragments that were smaller than 1 kb or bigger than 5 kb, DNA fragments of 1 - 5 kb were treated with End-It DNA End-Repair Kit (Epicentre, Madison, WI, USA) and subjected to phenol-chloroform extraction to eliminate any T4 polynucleokinase activity. Expression vector pET-30c (+) (Novagen, Madison, WI, USA) was digested with EcoRV, purified with QIAquick PCR Purification Kit (Qiagen, Valencia, CA, USA), and dephosphorylated with APex heat-labile alkaline phosphatase (Epicentre). A total of 250 ng of pET-30c (+)/EcoRV and blunt-ended E. faecalis DNA fragments at size of 1 - 5 kb were ligated at various molar ratios of vector to insert DNA (1:0.5, 1:1, 1:2, 1:5, and 1:10) using T4 DNA ligase (New England Biolab, Ipswich, MA, USA). The ligated mixture was used to transform E. coli DH5α (Takara, Daejon, Korea). Transformants were selected on LB plates containing kanamycin antibiotics. A total of 10 colonies were randomly selected and screened using colony PCR with T7 promoter primers. The frequency of self-ligation was lower than 20%. Purified recombinant DNA mixture was used to transform E. coli BL21 (DE3)/pLysS Competent Cells (Stratagene). Transformed cells were grown at 37℃ overnight on LB agar plates containing kanamycin (30 µg/mL).
Animal experimental protocols were reviewed and approved by the Institutional Animal Care and Use Committee, Chonnam National University (approval number: YB-2011-19). All surgical procedures were performed under sterile conditions. One 3 year old male beagle dog with weight of 10 kg was used for this study. Anesthetic induction was achieved by intravenous administration of thiopental (13.2 mg/kg body weight) followed by administration of 1 - 2% of isoflurane via an endotrachial tube. Additional local anesthesia of 2% lidocaine hydrochloride with 1:100,000 epinephrine was used to give a regional block. Preoperative radiograph was taken to determine any existing periodontal or endodontic lesion. After the induction, the second, third, and fourth premolars of each side of the upper jaw were used for inducing experimental endodontic infection as follows. A high-speed handpiece equipped with a round bur with saline irrigation was used to perforate a buccal surface of each tooth until it reached the pulp tissue. A total of 1.5 × 105
E. faecalis cells were then injected into the pulp cavity using an insulin syringe to induce endodontic infection. The access opening on the buccal surface was subsequently covered with light-cured composite materials. After the surgical procedure, analgesic medication (3 mg/kg body weight, Ketopro, Unibiotech Co., Yesan, Korea) was administered for three days.
Three weeks after injecting bacterial cells into the pulp, a radiograph was taken again to assess the status of periapical lesion of infected teeth. At this time, the dog was anesthetized again using the same protocol described above. All infected premolars were extracted and hemisectioned. Infected pulp and root canal tissues were removed using a barbed broach. They were immediately dissolved in phosphate buffered saline (PBS). Bacterial cell pellets were obtained by centrifugation at ×4,000 g for 20 minutes as described previously.17 Purified cell pellets were immediately stored in -20℃. They were sent to a commercial vendor (Young In Frontier Co., Seoul, Korea) to produce hyperimmune antisera in rabbits. Based on optical density, it was determined that the bacterial cell pellets contained more than 5 × 109 cells. The presence of E. faecalis was confirmed by visual inspection under a light microscope and sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) analysis. The efficacy of produced antibodies was monitored by the enzyme-linked immunosorbent assay (ELISA) analysis.
To obtain a serum probe that reacted only to in vivo expressed E. faecalis antigens, hyperimmune rabbit sera were adsorbed with E. faecalis cells cultivated in vitro. Adsorptions were performed using whole bacterial cells. First, rabbit polyclonal antiserum was mixed with 1 × 109
E. faecalis cells with slow agitation (30 rpm) at 4℃ for 1 hour. Cells were removed by centrifugation at ×5,000 g for 10 minutes and the serum (supernatant) was recovered. This step was repeated 4 times. After centrifugation, adsorbed serum was retrieved and stored at -80℃ until used. Cell lysates bound to nitrocellulose membrane were added into serum after removing whole cells and agitated overnight at 4℃. The serum was collected the next day. To remove antibodies that bound to E. coli proteins, the serum was agitated again with nitrocellulose membrane-bound E. coli BL21 harboring pET-30c(+) vector only.
To determine the adsorption efficiency, ELISA was performed as described previously.18 Briefly, a 96-well plate was coated with E. faecalis crude protein extracts at a concentration of 0.1 mg/100 mL in a coating buffer (0.1 M sodium carbonate, pH 9.6) overnight at 4℃. After washing, a blocking solution (2% skim milk in PBS, 300 µL) was added and incubated at 37℃ for 1 hour, followed by washing with PBS. The adsorbed rabbit antisera was added to each well using a serial dilution starting from 1:1,000 dilution and incubated at 37℃ for 2 hours. After the incubation, the plate was washed three times with 1X Tris buffered saline and Tween (TBS-Tween). Then, 50 mL of 1:10,000 diluted goat anti-human IgG conjugated with horse radish peroxidase (Abcam, Hanam, Korea) was added to each well and incubated at 37℃ for 1 hour. After washing the wells three times with TBS-Tween solution, TMB solution (GenDEPOT, Barker, TX, USA) was added to each well. After incubating at room temperature for 2 minutes, the reaction was stopped by adding 1 N H2SO4 (100 mL). Absorbance was measured at 450 nm using a 96-well plate reader (spectrometer).
E. coli recombinant clones were screened using the adsorbed hyperimmune rabbit sera as described previously.19 Briefly, kanamycin-resistant E. coli clones (approximately 200 per plate) were transferred to a nitrocellulose membrane. In vivo grown E. faecalis and E. coli containing pET-30c (+) were spotted onto the membrane as positive and negative control for screening, respectively. The membrane was then transferred (colony side up) to a LB agar plate containing 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) to induce protein expression. After incubation at 37℃ for 3 hours, the membrane was removed and the attached cells were lysed by exposure to chloroform vapor. The membrane was then blocked with 5% skim milk in PBS for 1 hour, washed with PBS-Tween 20, and incubated overnight with the adsorbed rabbit hyperimmune serum (1:500) at 4℃. After the incubation, the membrane was washed and probed with goat anti-rabbit IgG conjugated with horse radish peroxidase (1:1,000) at room temperature for 1 hour. A 1-step chloronaphthol (4CN, Thermo Scientifc, Waltham, MA, USA) was used as substrate to identify reactive clones. Positive clones showing reactivity were isolated from the original (master) plate. They were grown on LB-kanamycin plates. Their reactivity was confirmed again using a 4CN as substrate.
Plasmid DNAs from positive clones were purified using a QIAprep Spin Miniprep Kit (Qiagen, Valencia, CA, USA). DNA sequences of inserted DNA fragments from E. faecalis were determined by direct sequencing using a T7 promoter primer with an ABI Prism 377 automatic DNA sequencer by double-strand dideoxy chain termination method at GenoTech Corp. (Daejeon, Korea). Identified gene sequences were compared to DNA and protein databases using a BLAST program (http://www.ncbi.nlm.nih.gov) and analyzed by Vector NTI Software (Invitrogen, Carlsbad, CA, USA). Functional classification of identified antigens was based on published studies of identified proteins of E. faecalis if available.
Results
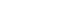
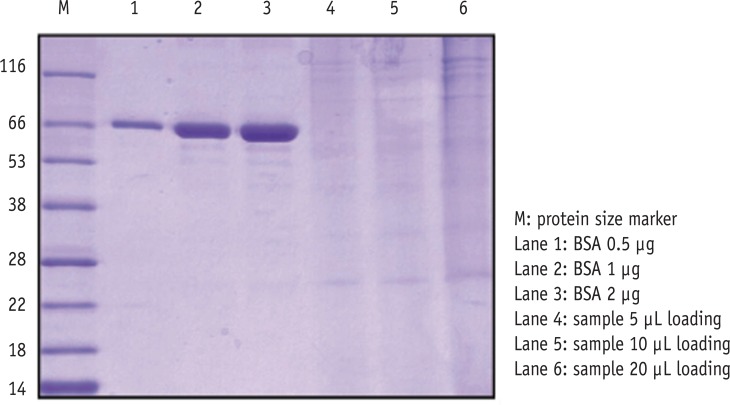
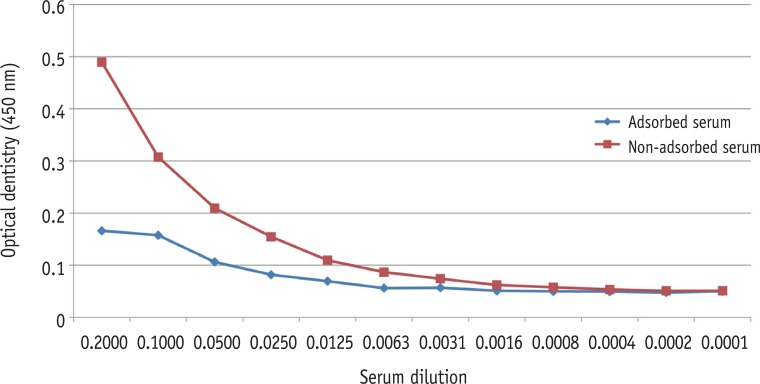
SDS-PAGE analysis confirmed the presence of bacterial cells in the sample acquired from experimental endodontic infection (Figure 1). The hyperimmune rabbit antisera produced against E. faecalis infected pulp and periapical tissues were successively adsorbed against E. faecalis cells grown in vitro. The adsorption efficiency was determined by examining immuno-reactivity of antisera using ELISA after each adsorption step with in vitro grown E. faecalis. Fully adsorbed serum exhibited a significant lower reactivity to in vitro grown E. faecalis (Figure 2), suggesting that the adsorption process efficiently removed antibodies against most E. faecalis proteins produced during in vitro cultivation.
Plasmid DNAs of ten colonies randomly selected from the library were subjected to direct DNA sequencing to confirm the presence of the DNA inserts present in the recombinant plasmids. Of the 10 colonies, eight had DNA inserts in the pET-30c (+) vector. A total of approximately 4,500 recombinant clones were screened using the adsorbed pooled serum. In vivo grown E. faecalis cells and E. coli cells containing only pET-30c (+) placed on the nitrocellulose membrane was used as positive and negative control, respectively, for the immunoscreening. Initial screening resulted in the detection of 30 immuno-reactive clones. Of the 30 clones, nineteen reproducibly exhibited reactivity with the adsorbed serum. Therefore, they were subjected to further DNA sequencing.
Sequencing of the DNA inserts of the 19 positive clones revealed 16 open reading frames (ORFs). Sequences of the remaining three clones could not be obtained. Sequences of the 16 clones were analyzed using BLAST program. CMAT-identified in vivo-induced genes and their predicted functions are listed in Table 1. Many determined gene sequences contained portions of ORFs for putative genes. The size of ORF varied from 663 bp to 2,911 bp. In addition, multiple ORFs were not identified. Sequence analysis revealed that many of these ORFs were enzymes implicated in housekeeping functions with intermediary metabolism. In addition, conserved hypothetical proteins with unknown function and putative membrane protein were also identified, along with copper resistance proteins.
Discussion
Very limited information regarding the nature of virulence factors of E. faecalis has hampered our understanding of the mechanisms by which this endodontic pathogen interacts with the host and ultimately leads to endodontic infection. Considering that genes (proteins) expressed in vivo are very likely to be important in the microorganisms' ability to survive in the host and/or serve as virulence factors, our major objective was to identify E. faecalis proteins that were uniquely expressed in vivo during the actual infectious process. For this purpose, we adopted a new technique called CMAT designed to detect immunoreactive proteins exhibiting specificity only to in vivo-induced proteins. For this immunoscreening, we needed to prepare a 'probe' antibody that reacts only to in vivo-induced proteins (not proteins produced in vitro). CMAT is a modification of the IVIAT that has enabled many investigators to identify in vivo-induced proteins directly from the human host rather than from a potentially misleading animal model.15,19 IVIAT has been successfully used to study pathogens such as Candida albicans, Vibrio cholera, Mycobacterium tuberculosis, Vibrio vulnificus, Escherichia coli, Salmonella enterica, Group A Streptococcus, and periodontal pathogens including Aggregatibacter actinomycetemcomitan, Porphyromonas gingivalis, and Tannerella forsythia.18,20,21,22,23,24,25,26,27,28 However, the use of IVIAT requires high antibody titers in human sera against the etiological agents, it may not be readily applicable to the study of microorganisms that do not induce strong humoral responses in humans. Therefore, CMAT was specifically developed to address the above limitation of IVIAT.
The original design of this study was to use E. faecalis samples from clinical cases of pulp and endodontic infection from human subjects. However, we failed to secure adequate amount of E. faecalis cells from clinical cases that could induce sufficient humoral immune responses (data not shown). Therefore, as an alternative, we adopted an animal model using a beagle dog for the purpose of producing in vivo-grown E. faecalis cells. In this model, pulp tissues were intentionally infected with E. faecalis cells in hopes of causing endodontic infection. From the lesions of pulp and root canal infections, we were able to collect adequate amount of E. faecalis cells for the CMAT purpose.
In this pilot study, we used CMAT to identify more than a dozen protein antigens that were reactive with adsorbed antisera reactive with in vivo-grown E. faecalis cells. To the best of our knowledge, this is the first attempt to identify in vivo-induced proteins from E. faecalis. One of the advantages of using CMAT is that it does not require an efficient genetic manipulation system nor human infection because it can utilize an alternative animal model.
Many in vivo-induced genes of E. faecalis identified by CMAT included several enzymes related to housekeeping functions, including energy metabolism, translation, amino acid biosynthesis, and other cellular process (Table 1). These genes were similar to those identified by previous IVIAT studies.18,29 These genes might play roles that are not required for in vitro growth.30 It has been observed that 'housekeeping' genes are not constitutively expressed at the transcriptional level in bacteria as previously assumed, indicating that housekeeping metabolism as a very dynamic process is extremely capable of adapting to different growth conditions.31,32,33 Therefore, it is plausible that these genes are up-regulated and expressed only in vivo.
A copper resistance protein was identified in this study. Although its exact function is unknown, it has been found in other bacteria including E. coli, Pseudomonas syringae, and Xanthomonas axonopodis.34 Considering metallic copper can act as antimicrobial surface, copper resistance could function as a potential virulence factor.35 CMAT was able to identify several putative membrane proteins. Since little is known regarding the functions of membrane protein in E. faecalis, analysis of these in vivo induced proteins identified in this study may increase our understanding on the pathogenic mechanisms of this microorganism.
Further use of CMAT in E. faecalis will be required to identify additional in vivo induced proteins that are likely to be important in survival in the host and may serve as virulence factors in the disease process. In future studies, the functional roles of these identified genes can be tested by constructing isogenic mutants for use in animal models or in vitro models. These efforts will ultimately allow us to gain a better insight into the pathogenic mechanisms of E. faecalis. For example, gene expression (mRNA) analysis using real-time PCR for each identified gene could be performed to confirm its in vivo expression.18,36 Genetic analysis using isogenic mutant (such as siRNA-knockdown or gene destruction) could be performed to confirm the identified genes (proteins) are really related to the disease process, as previously performed.37,38
Once functional roles are confirmed to be important for E. faecalis virulence, these virulence factors could be analyzed as candidates for diagnostic, therapeutic, and preventive measures for endodontic infections. Identified virulence factors could be used as disease markers for pulpal and peri-apical infection associated with E. faecalis. The presence of these virulence factors in the pulp and periaplical tissues will confirm the infectious disease process mediated by E. faecalis without bacterial culture of the clinical samples. Another potential application includes the development of therapeutic and preventive approaches targeting these virulence factors, such as intracanal medication, vaccines, and novel antimicrobial substances.22 By specifically inactivating virulence factors without affecting other non-virulent components, it would be possible to effectively inhibit or reduce virulence mediated by E. faecalis. These efforts will lead to improved management of persistent endodontic infections associated with E. faecalis.
Conclusions
Sixteen in vivo-induced proteins of E. faecalis were identified using CMAT and experimental endodontic infection model. These proteins were expressed uniquely in vivo and reactive with sera produced against in vivo-grown E. faecalis. The results of this pilot study suggest that CMAT adopting an experimental animal model of endodontic infection is a useful approach to identify and characterize potential virulence factors of E. faecalis. Detailed analysis of these in vivo-induced genes (proteins) will lead to a better understanding of the molecular mechanisms involved in the endodontic infection of E. faecalis.
Acknowledgment
This research was supported in part by Korean National Research Foundation (2010-0002871) and Chonnam National University Hospital Research Institute of Clinical Medicine (CRI 12020-1).
- 1. Finlay BB, Falkow S. Common themes in microbial pathogenicity revisited. Microbiol Mol Biol Rev 1997;61:136-169.ArticlePubMedPMCPDF
- 2. Falkow S. Molecular Koch's postulates applied to microbial pathogenicity. Rev Infect Dis 1988;10(Suppl 2):S274-S276.ArticlePubMed
- 3. Sundqvist G, Johansson E, Sjögren U. Prevalence of black-pigmented bacteroides species in root canal infections. J Endod 1989;15:13-19.ArticlePubMed
- 4. Griffee MB, Patterson SS, Miller CH, Kafrawy AH, Newton CW. The relationship of Bacteroides melaninogenicus to symptoms associated with pulpal necrosis. Oral Surg Oral Med Oral Pathol 1980;50:457-461.ArticlePubMed
- 5. Sundqvist GK, Eckerbom MI, Larsson AP, Sjögren UT. Capacity of anaerobic bacteria from necrotic dental pulps to induce purulent infections. Infect Immun 1979;25:685-693.ArticlePubMedPMCPDF
- 6. Sundqvist G, Figdor D, Persson S, Sjögren U. Microbiologic analysis of teeth with failed endodontic treatment and the outcome of conservative retreatment. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 1998;85:86-93.ArticlePubMed
- 7. Hancock HH 3rd, Sigurdsson A, Trope M, Moiseiwitsch J. Bacteria isolated after unsuccessful endodontic treatment in a North American population. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2001;91:579-586.ArticlePubMed
- 8. Kayaoglu G, Ørstavik D. Virulence factors of Enterococcus faecalis: relationship to endodontic disease. Crit Rev Oral Biol Med 2004;15:308-320.PubMed
- 9. Smith H. Questions about the behaviour of bacterial pathogens in vivo. Philos Trans R Soc Lond B Biol Sci 2000;355:551-564.ArticlePubMedPMCPDF
- 10. Angelichio MJ, Camilli A. In vivo expression technology. Infect Immun 2002;70:6518-6523.ArticlePubMedPMCPDF
- 11. Mahan MJ, Slauch JM, Hanna PC, Camilli A, Tobias JW, Waldor MK, Mekalanos JJ. Selection for bacterial genes that are specifically induced in host tissues: the hunt for virulence factors. Infect Agents Dis 1993;2:263-268.PubMed
- 12. Hensel M, Shea JE, Gleeson C, Jones MD, Dalton E, Holden DW. Simultaneous identification of bacterial virulence genes by negative selection. Science 1995;269:400-403.ArticlePubMed
- 13. Valdivia RH, Falkow S. Fluorescence-based isolation of bacterial genes expressed within host cells. Science 1997;277:2007-2011.ArticlePubMed
- 14. Ramachandran N, Raphael JV, Hainsworth E, Demirkan G, Fuentes MG, Rolfs A, Hu Y, LaBaer J. Next-generation high-density self-assembling functional protein arrays. Nat Methods 2008;5:535-538.ArticlePubMedPMCPDF
- 15. Rollins SM, Peppercorn A, Hang L, Hillman JD, Calderwood SB, Handfield M, Ryan ET. In vivo induced antigen technology (IVIAT). Cell Microbiol 2005;7:1-9.ArticlePubMed
- 16. Handfield M, Hillman JD. In vivo induced antigen technology (IVIAT) and change mediated antigen technology (CMAT). Infect Disord Drug Targets 2006;6:327-334.ArticlePubMed
- 17. Lee SW, Hillman JD, Progulske-Fox A. The hemagglutinin genes hagB and hagC of Porphyromonas gingivalis are transcribed in vivo as shown by use of a new expression vector. Infect Immun 1996;64:4802-4810.ArticlePubMedPMCPDF
- 18. Yoo JY, Kim HC, Zhu W, Kim SM, Sabet M, Handfield M, Hillman J, Progulske-Fox A, Lee SW. Identification of Tannerella forsythia antigens specifically expressed in patients with periodontal disease. FEMS Microbiol Lett 2007;275:344-352.ArticlePubMed
- 19. Handfield M, Brady LJ, Progulske-Fox A, Hillman JD. IVIAT: a novel method to identify microbial genes expressed specifically during human infections. Trends Microbiol 2000;8:336-339.ArticlePubMed
- 20. Cheng S, Clancy CJ, Checkley MA, Handfield M, Hillman JD, Progulske-Fox A, Lewin AS, Fidel PL, Nguyen MH. Identification of Candida albicans genes induced during thrush offers insight into pathogenesis. Mol Microbiol 2003;48:1275-1288.ArticlePubMed
- 21. Hang L, John M, Asaduzzaman M, Bridges EA, Vanderspurt C, Kirn TJ, Taylor RK, Hillman JD, Progulske-Fox A, Handfield M, Ryan ET, Calderwood SB. Use of in vivo-induced antigen technology (IVIAT) to identify genes uniquely expressed during human infection with Vibrio cholerae. Proc Natl Acad Sci U S A 2003;100:8508-8513.ArticlePubMedPMC
- 22. Deb DK, Dahiya P, Srivastava KK, Srivastava R, Srivastava BS. Selective identification of new therapeutic targets of Mycobacterium tuberculosis by IVIAT approach. Tuberculosis (Edinb) 2002;82:175-182.ArticlePubMed
- 23. Kim YR, Lee SE, Kim CM, Kim SY, Shin EK, Shin DH, Chung SS, Choy HE, Progulske-Fox A, Hillman JD, Handfield M, Rhee JH. Characterization and pathogenic significance of Vibrio vulnificus antigens preferentially expressed in septicemic patients. Infect Immun 2003;71:5461-5471.ArticlePubMedPMCPDF
- 24. John M, Kudva IT, Griffin RW, Dodson AW, McManus B, Krastins B, Sarracino D, Progulske-Fox A, Hillman JD, Handfield M, Tarr PI, Calderwood SB. Use of in vivo-induced antigen technology for identification of Escherichia coli O157:H7 proteins expressed during human infection. Infect Immun 2005;73:2665-2679.PubMedPMC
- 25. Harris JB, Baresch-Bernal A, Rollins SM, Alam A, LaRocque RC, Bikowski M, Peppercorn AF, Handfield M, Hillman JD, Qadri F, Calderwood SB, Hohmann E, Breiman RF, Brooks WA, Ryan ET. Identification of in vivo-induced bacterial protein antigens during human infection with Salmonella enterica serovar Typhi. Infect Immun 2006;74:5161-5168.ArticlePubMedPMCPDF
- 26. Salim KY, Cvitkovitch DG, Chang P, Bast DJ, Handfield M, Hillman JD, de Azavedo JC. Identification of group A Streptococcus antigenic determinants upregulated in vivo. Infect Immun 2005;73:6026-6038.ArticlePubMedPMCPDF
- 27. Cao SL, Progulske-Fox A, Hillman JD, Handfield M. in vivo induced antigenic determinants of Actinobacillus actinomycetemcomitans. FEMS Microbiol Lett 2004;237:97-103.ArticlePubMed
- 28. Song YH, Kozarov EV, Walters SM, Cao SL, Handfield M, Hillman JD, Progulske-Fox A. Genes of periodontopathogens expressed during human disease. Ann Periodontol 2002;7:38-42.ArticlePubMed
- 29. Gu H, Zhu H, Lu C. Use of in vivo-induced antigen technology (IVIAT) for the identification of Streptococcus suis serotype 2 in vivo-induced bacterial protein antigens. BMC Microbiol 2009;9:201.ArticlePubMedPMCPDF
- 30. Lee HR, Rhyu IC, Kim HD, Jun HK, Min BM, Lee SH, Choi BK. In vivo-induced antigenic determinants of Fusobacterium nucleatum subsp. nucleatum. Mol Oral Microbiol 2011;26:164-172.ArticlePubMed
- 31. Savli H, Karadenizli A, Kolayli F, Gundes S, Ozbek U, Vahaboglu H. Expression stability of six housekeeping genes: A proposal for resistance gene quantification studies of Pseudomonas aeruginosa by real-time quantitative RT-PCR. J Med Microbiol 2003;52:403-408.ArticlePubMed
- 32. Vandecasteele SJ, Peetermans WE, Merckx R, Van Eldere J. Quantification of expression of Staphylococcus epidermidis housekeeping genes with Taqman quantitative PCR during in vitro growth and under different conditions. J Bacteriol 2001;183:7094-7101.ArticlePubMedPMCPDF
- 33. Widada J, Nojiri H, Kasuga K, Yoshida T, Habe H, Omori T. Quantification of the carbazole 1,9a-dioxygenase gene by real-time competitive PCR combined with coextraction of internal standards. FEMS Microbiol Lett 2001;202:51-57.ArticlePubMed
- 34. Espirito Santo C, Taudte N, Nies DH, Grass G. Contribution of copper ion resistance to survival of Escherichia coli on metallic copper surfaces. Appl Environ Microbiol 2008;74:977-986.ArticlePubMedPDF
- 35. Grass G, Rensing C, Solioz M. Metallic copper as an antimicrobial surface. Appl Environ Microbiol 2011;77:1541-1547.ArticlePubMedPDF
- 36. Walters S, Belanger M, Rodrigues PH, Whitlock J, Progulske-Fox A. A member of the peptidase M48 superfamily of Porphyromonas gingivalis is associated with virulence in vitro and in vivo. J Oral Microbiol 2009;1.
- 37. Yuan L, Rodrigues PH, Bélanger M, Dunn W Jr, Progulske-Fox A. The Porphyromonas gingivalis clpB gene is involved in cellular invasion in vitro and virulence in vivo. FEMS Immunol Med Microbiol 2007;51:388-398.ArticlePubMed
- 38. Yuan L, Rodrigues PH, Bélanger M, Dunn WA Jr, Progulske-Fox A. Porphyromonas gingivalis htrA is involved in cellular invasion and in vivo survival. Microbiology 2008;154:1161-1169.ArticlePubMed
REFERENCES
Figure 1
Confirmation of bacterial antigens by SDS-PAGE. In lanes 4 - 6, different amount of bacterial cell pellets were loaded. SDS-PAGE, sodium dodecyl sulphate-polyacrylamide gel electrophoresis; BSA, bovine serum albumin.
Figure 2
Testing an efficacy of the adsorption procedure for hyperimmune rabbit antisera against E. faecalis whole cell pellets by ELISA analysis. Adsorbed serum exhibited a significant decrease of the reactivity. ELISA, enzyme-linked immunosorbent assay.
Table 1
E. faecalis protein antigens identified by CMAT
Tables & Figures
REFERENCES
Citations
Citations to this article as recorded by 
-
Insights into ecology, pathogenesis, and biofilm formation of
Enterococcus faecalis
from functional genomics
Julia L. E. Willett, Gary M. Dunny, Corrella S. Detweiler
Microbiology and Molecular Biology Reviews.2025;[Epub] CrossRef - Antibody-based Screening of Porphyromonas gingivalis Proteins Specifically Produced in Patients with Chronic Periodontitis
Hye-Jung Kim, Seok-Woo Lee
International Journal of Oral Biology.2018; 43(4): 201. CrossRef - The controversial role of Enterococcus faecalis in colorectal cancer
Carolina Vieira de Almeida, Antonio Taddei, Amedeo Amedei
Therapeutic Advances in Gastroenterology.2018;[Epub] CrossRef - Model systems for the study of Enterococcal colonization and infection
H. M. Sharon Goh, M. H. Adeline Yong, Kelvin Kian Long Chong, Kimberly A. Kline
Virulence.2017; 8(8): 1525. CrossRef
 ePub Link
ePub Link Cite
CiteIdentification of Enterococcus faecalis antigens specifically expressed in vivo
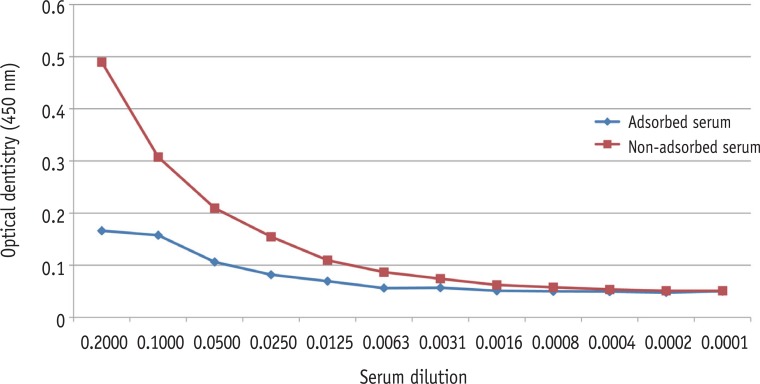
Figure 1 Confirmation of bacterial antigens by SDS-PAGE. In lanes 4 - 6, different amount of bacterial cell pellets were loaded. SDS-PAGE, sodium dodecyl sulphate-polyacrylamide gel electrophoresis; BSA, bovine serum albumin.
Figure 2 Testing an efficacy of the adsorption procedure for hyperimmune rabbit antisera against E. faecalis whole cell pellets by ELISA analysis. Adsorbed serum exhibited a significant decrease of the reactivity. ELISA, enzyme-linked immunosorbent assay.
Figure 1
Figure 2
Identification of Enterococcus faecalis antigens specifically expressed in vivo
E. faecalis protein antigens identified by CMAT
| GenBank Protein ID | Protein name | Function |
|---|---|---|
| AFA92809.1 | Deoxyribose-phosphate aldolase | Energy metabolism |
| AEA94635.1 | Selenide, water dikinase | Energy metabolism |
| ADX80204.1 | Beta-glucosidase | Energy metabolism |
| AA079953.1 | Oxidoreductase | Energy metabolism |
| PF08503.3 | Tetrahydrodipicolinate N-acetyltransferase | Energy metabolism |
| ADX80368.1 | 3-ketoacyl-(acyl-carrier-protein) reductase | Energy metabolism |
| AEA93036.1 | Diaminopimelate epimerase | Energy metabolism |
| AEA92693.1 | DNA topoisomerase subunit A | Transcription |
| EF62_0348 | rRNA-23S ribosomal RNA | Transcription |
| AEA93730.1 | Translation initiation factor IF2 | Transcription |
| ADX79467.1 | Alanine racemase | Amino acid biosynthesis |
| AEA95149.1 | Response regulator sensor histidine kinase | Cellular process |
| ACF94631.1 | Copper resistance protein | Energy metabolism |
| AA080103.1 | Putative membrane protein | Outer membrane protein |
| ADX79209.1 | Conserved hypothetical protein | Unknown |
| ADX81283 | Conserved hypothetical protein | Unknown |
Table 1 E. faecalis protein antigens identified by CMAT